

Deriving Dorsal Spinal Sensory Interneurons from Human Pluripotent Stem Cells
- PMID: 29337120
- PMCID: PMC5832443
- DOI: 10.1016/j.stemcr.2017.12.012
Deriving Dorsal Spinal Sensory Interneurons from Human Pluripotent Stem Cells
Abstract
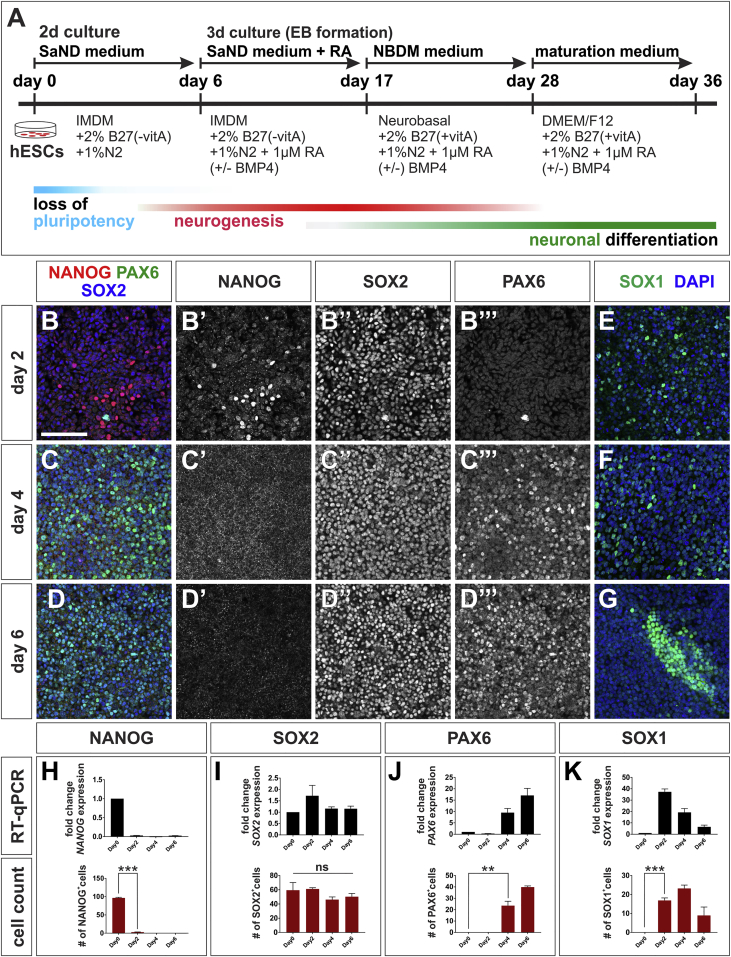
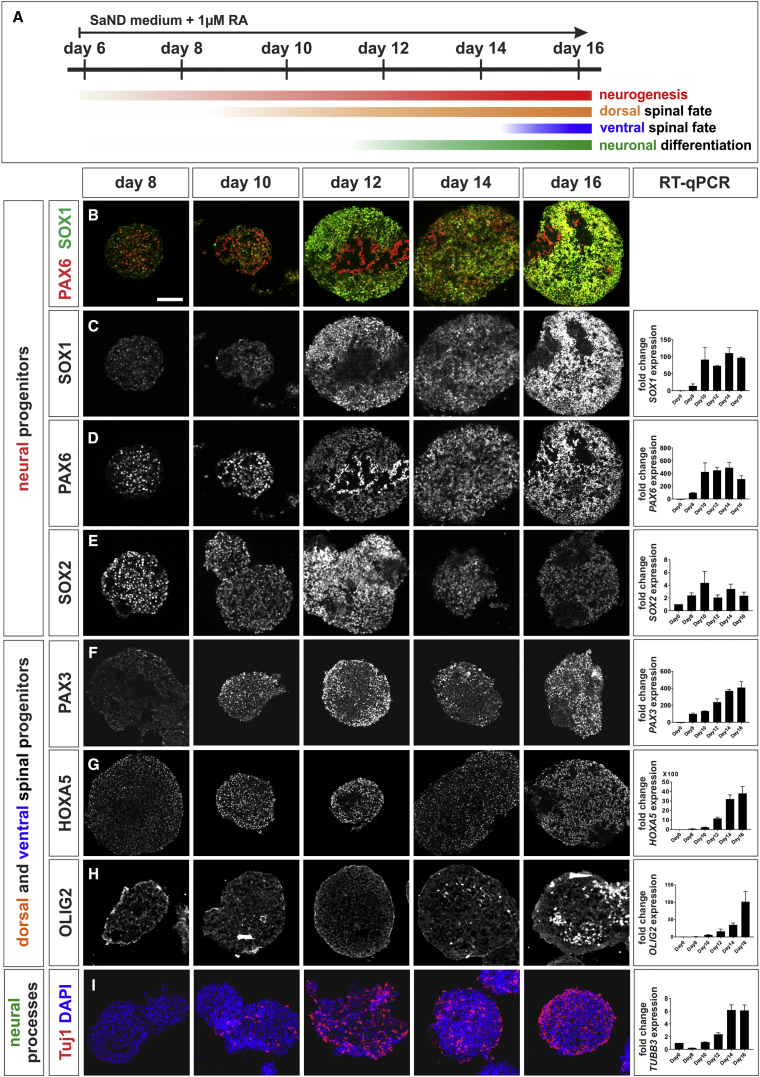
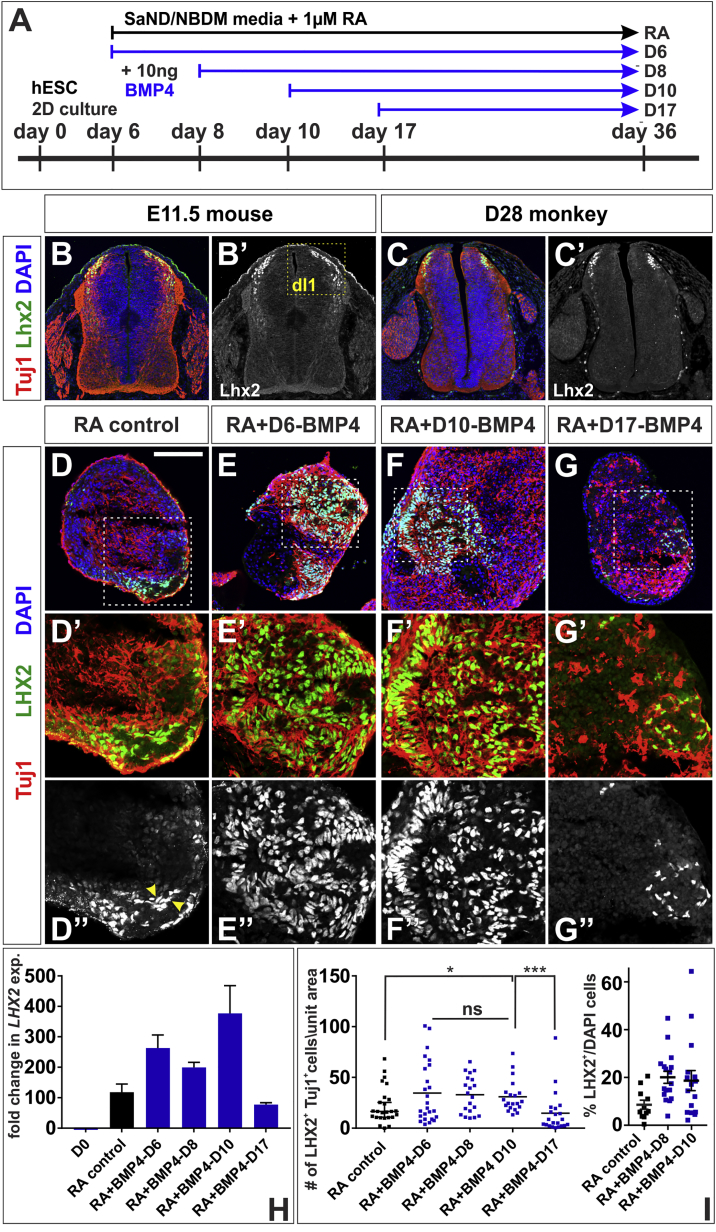
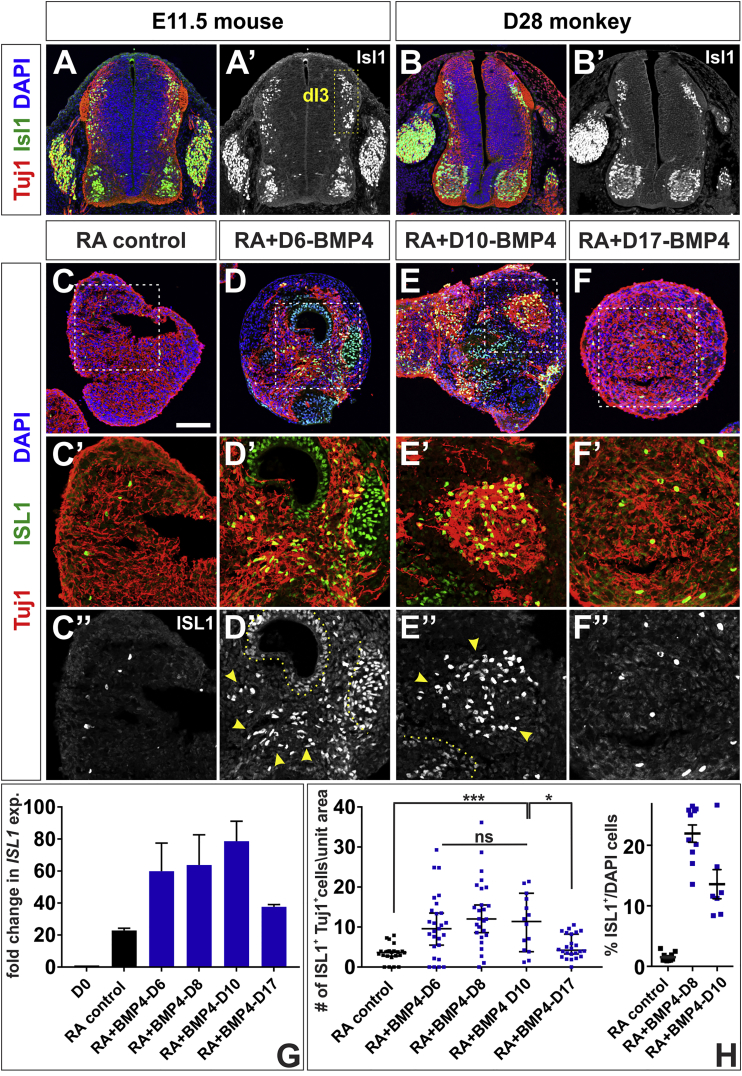
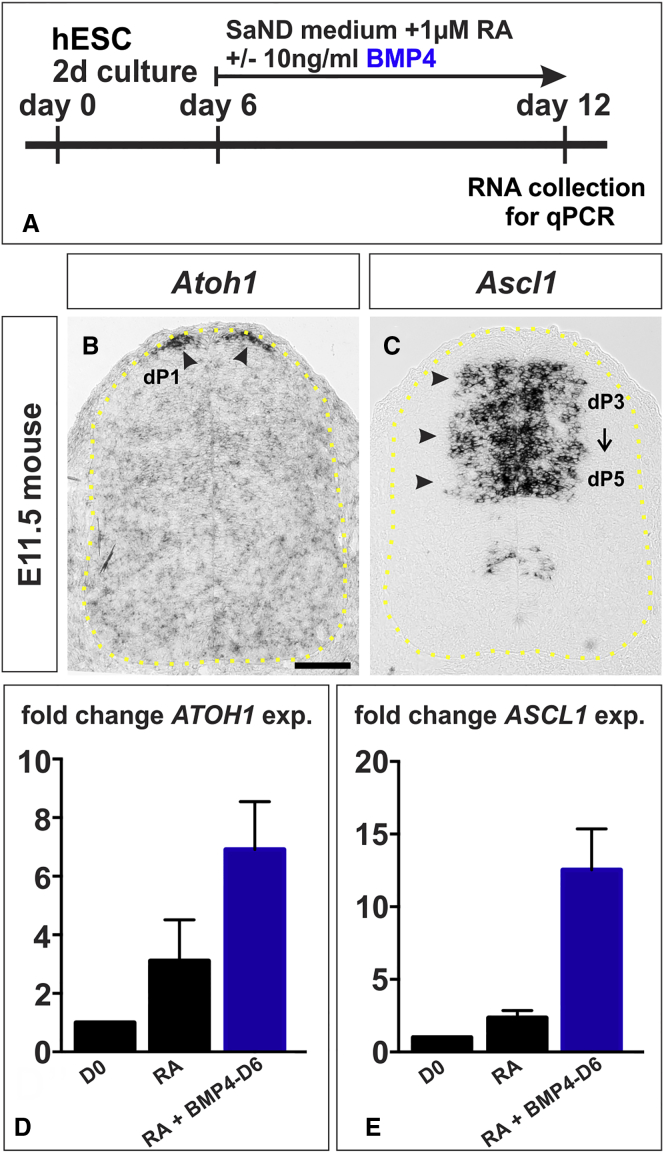
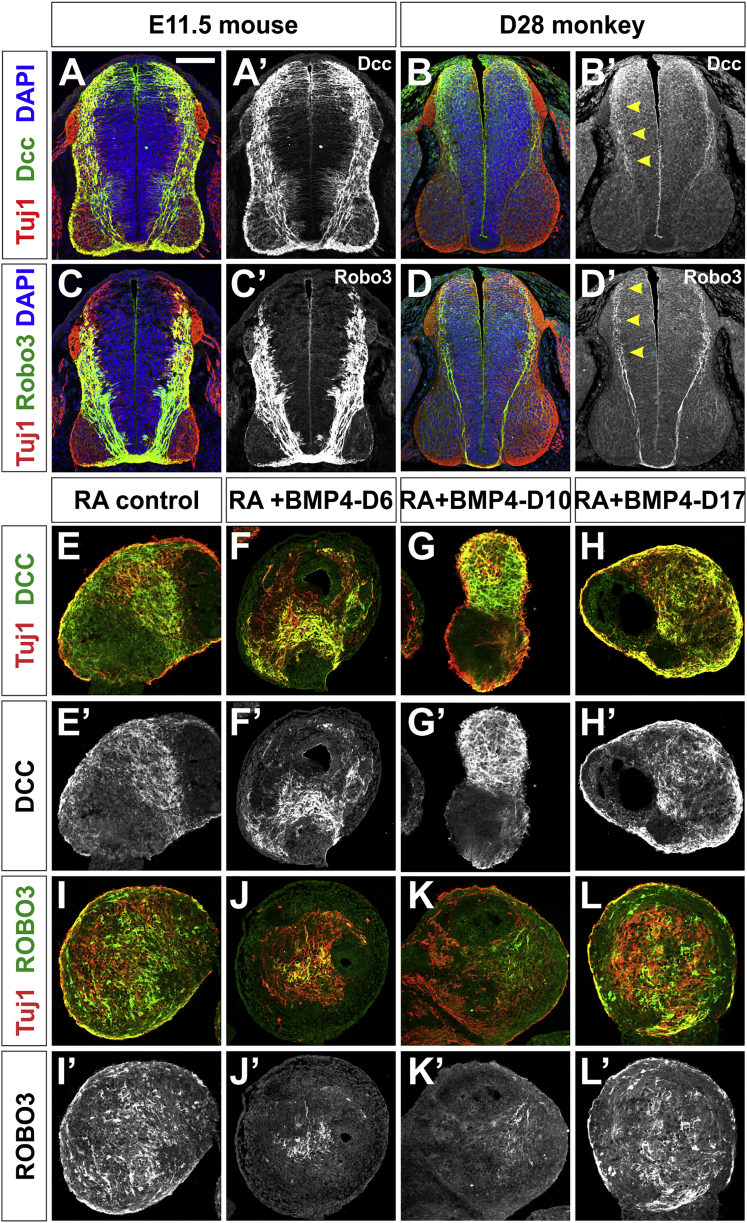
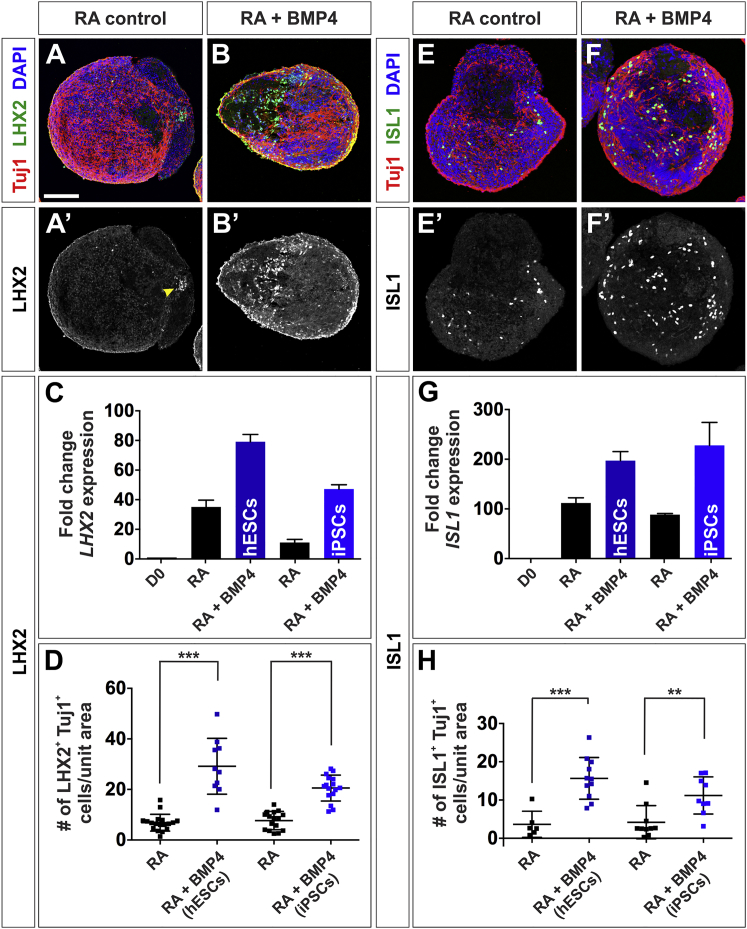
Cellular replacement therapies for neurological conditions use human embryonic stem cell (hESC)- or induced pluripotent stem cell (hiPSC)-derived neurons to replace damaged or diseased populations of neurons. For the spinal cord, significant progress has been made generating the in-vitro-derived motor neurons required to restore coordinated movement. However, there is as yet no protocol to generate in-vitro-derived sensory interneurons (INs), which permit perception of the environment. Here, we report on the development of a directed differentiation protocol to derive sensory INs for both hESCs and hiPSCs. Two developmentally relevant factors, retinoic acid in combination with bone morphogenetic protein 4, can be used to generate three classes of sensory INs: the proprioceptive dI1s, the dI2s, and mechanosensory dI3s. Critical to this protocol is the competence state of the neural progenitors, which changes over time. This protocol will facilitate developing cellular replacement therapies to reestablish sensory connections in injured patients.
Keywords: directed differentiation; human embryonic stem cells; induced pluripotent stem cells; mechanosensation; mouse spinal cord; neurons; primate spinal cord; proprioception; sensory interneurons; spinal cord.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Arber S., Han B., Mendelsohn M., Smith M., Jessell T.M., Sockanathan S. Requirement for the homeobox gene Hb9 in the consolidation of motor neuron identity. Neuron. 1999;23:659–674. - PubMed
-
- Bermingham N.A., Hassan B.A., Wang V.Y., Fernandez M., Banfi S., Bellen H.J., Fritzsch B., Zoghbi H.Y. Proprioceptor pathway development is dependent on Math1. Neuron. 2001;30:411–422. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
