

Immune Protection against Lethal Fungal-Bacterial Intra-Abdominal Infections
- PMID: 29339423
- PMCID: PMC5770546
- DOI: 10.1128/mBio.01472-17
Immune Protection against Lethal Fungal-Bacterial Intra-Abdominal Infections
Abstract
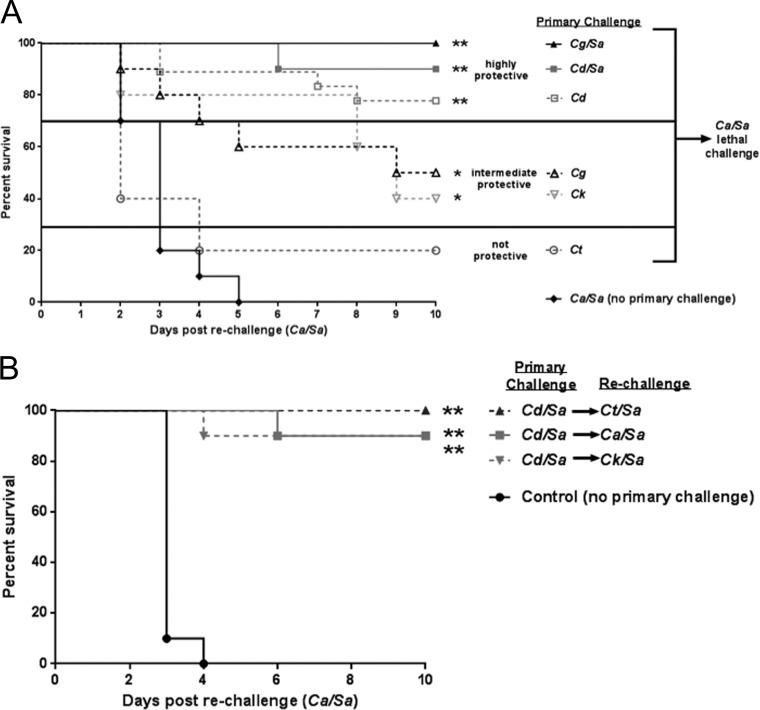
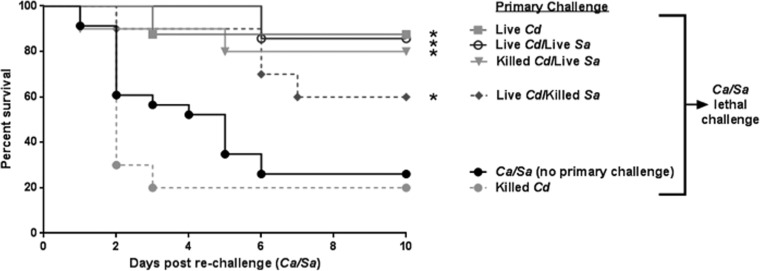
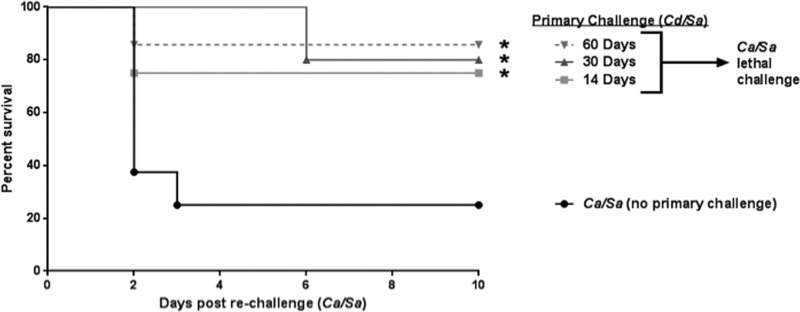
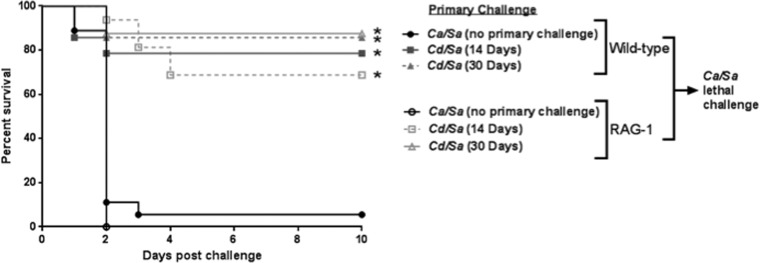
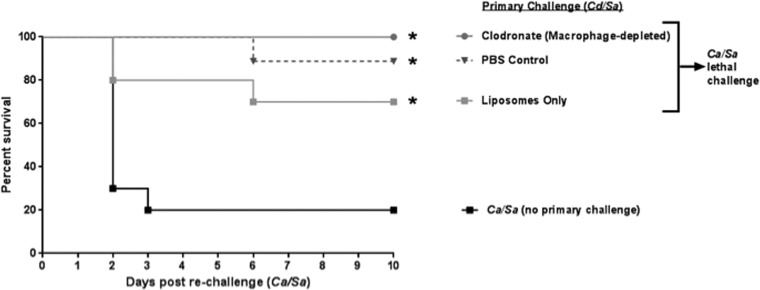
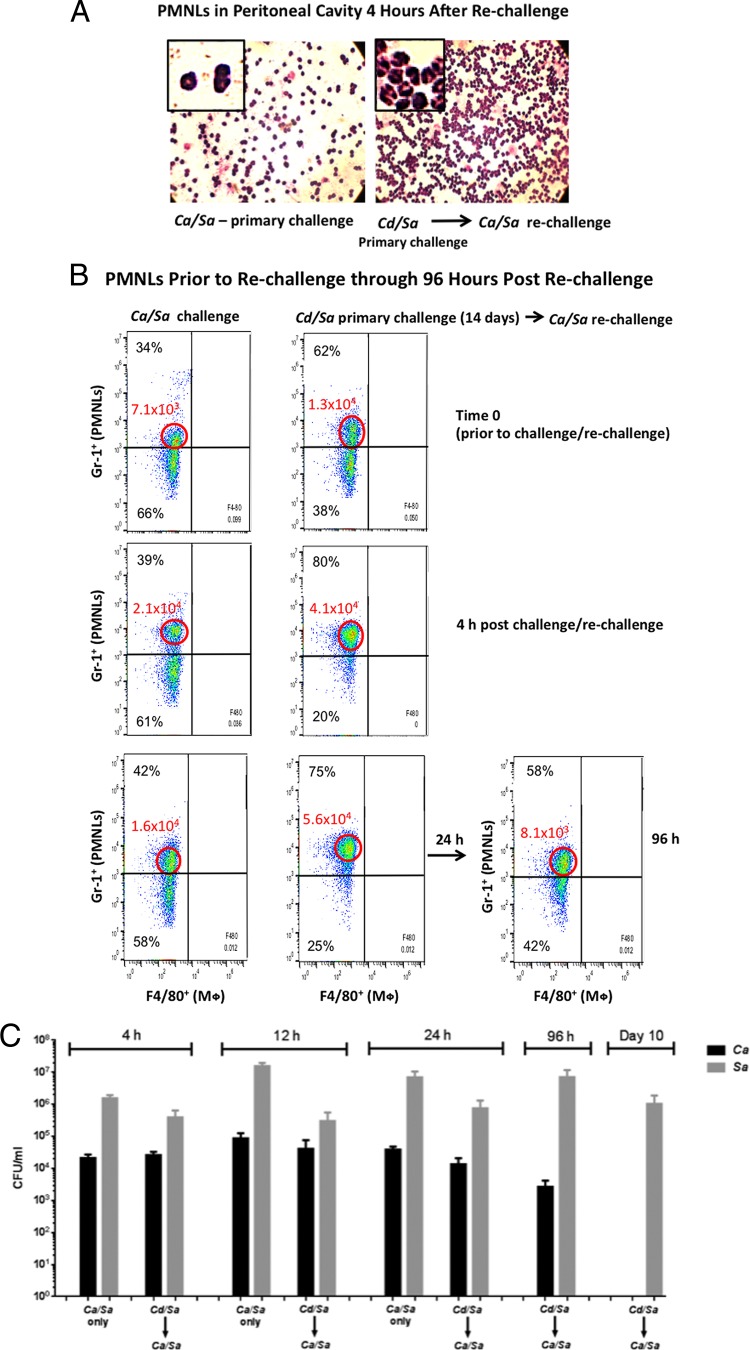
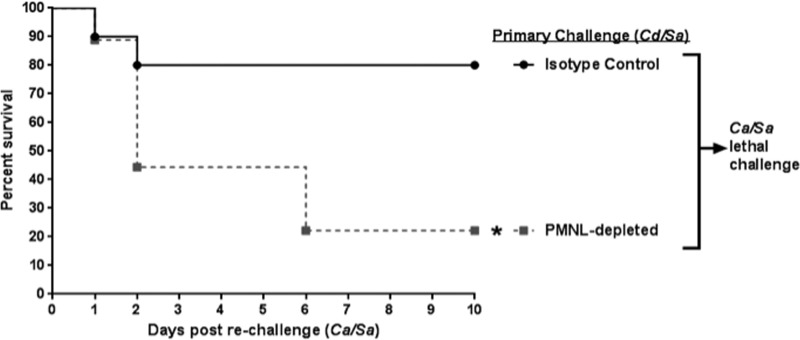
Polymicrobial intra-abdominal infections (IAIs) are clinically prevalent and cause significant morbidity and mortality, especially those involving fungi. Our laboratory developed a mouse model of IAI and demonstrated that intraperitoneal inoculation with Candida albicans or other virulent non-albicans Candida (NAC) species plus Staphylococcus aureus resulted in 70 to 80% mortality in 48 to 72 h due to robust local and systemic inflammation (sepsis). Surprisingly, inoculation with Candida dubliniensis or Candida glabrata with S. aureus resulted in minimal mortality, and rechallenge of these mice with lethal C. albicans/S. aureus (i.e., coninfection) resulted in >90% protection. The purpose of this study was to define requirements for C. dubliniensis/S. aureus-mediated protection and interrogate the mechanism of the protective response. Protection was conferred by C. dubliniensis alone or by killed C. dubliniensis plus live S. aureusS. aureus alone was not protective, and killed S. aureus compromised C. dubliniensis-induced protection. C. dubliniensis/S. aureus also protected against lethal challenge by NAC plus S. aureus and could protect for a long-term duration (60 days between primary challenge and C. albicans/S. aureus rechallenge). Unexpectedly, mice deficient in T and B cells (Rag-1 knockouts [KO]) survived both the initial C. dubliniensis/S. aureus challenge and the C. albicans/S. aureus rechallenge, indicating that adaptive immunity did not play a role. Similarly, mice depleted of macrophages prior to rechallenge were also protected. In contrast, protection was associated with high numbers of Gr-1hi polymorphonuclear leukocytes (PMNLs) in peritoneal lavage fluid within 4 h of rechallenge, and in vivo depletion of Gr-1+ cells prior to rechallenge abrogated protection. These results suggest that Candida species can induce protection against a lethal C. albicans/S. aureus IAI that is mediated by PMNLs and postulated to be a unique form of trained innate immunity.IMPORTANCE Polymicrobial intra-abdominal infections are clinically devastating infections with high mortality rates, particularly those involving fungal pathogens, including Candida species. Even in patients receiving aggressive antimicrobial therapy, mortality rates remain unacceptably high. There are no available vaccines against IAI, which is complicated by the polymicrobial nature of the infection. IAI leads to lethal systemic inflammation (sepsis), which is difficult to target pharmacologically, as components of the inflammatory response are also needed to control the infection. Our studies demonstrate that prior inoculation with low-virulence Candida species provides strong protection against subsequent lethal infection with C. albicans and S. aureus Surprisingly, protection is long-lived but not mediated by adaptive (specific) immunity. Instead, protection is dependent on cells of the innate immune system (nonspecific immunity) and provides protection against other virulent Candida species. This discovery implies that a form of trained innate immunity may be clinically effective against polymicrobial IAI.
Keywords: Candida albicans; Staphylococcus aureus; immune protection; innate immunity; intra-abdominal infection.
Copyright © 2018 Lilly et al.
Figures
References
-
- Heemken R, Gandawidjaja L, Hau T. 1997. Peritonitis: pathophysiology and local defense mechanisms. Hepatogastroenterology 44:927–936. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
