

Ontogeny of human mucosal-associated invariant T cells and related T cell subsets
- PMID: 29339446
- PMCID: PMC5789419
- DOI: 10.1084/jem.20171739
Ontogeny of human mucosal-associated invariant T cells and related T cell subsets
Abstract
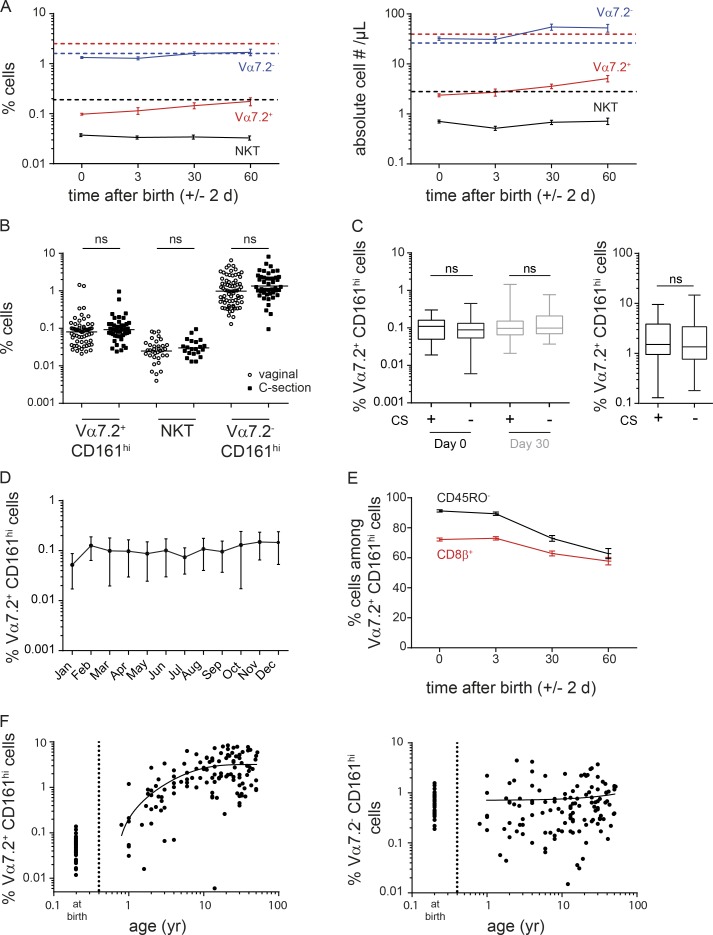
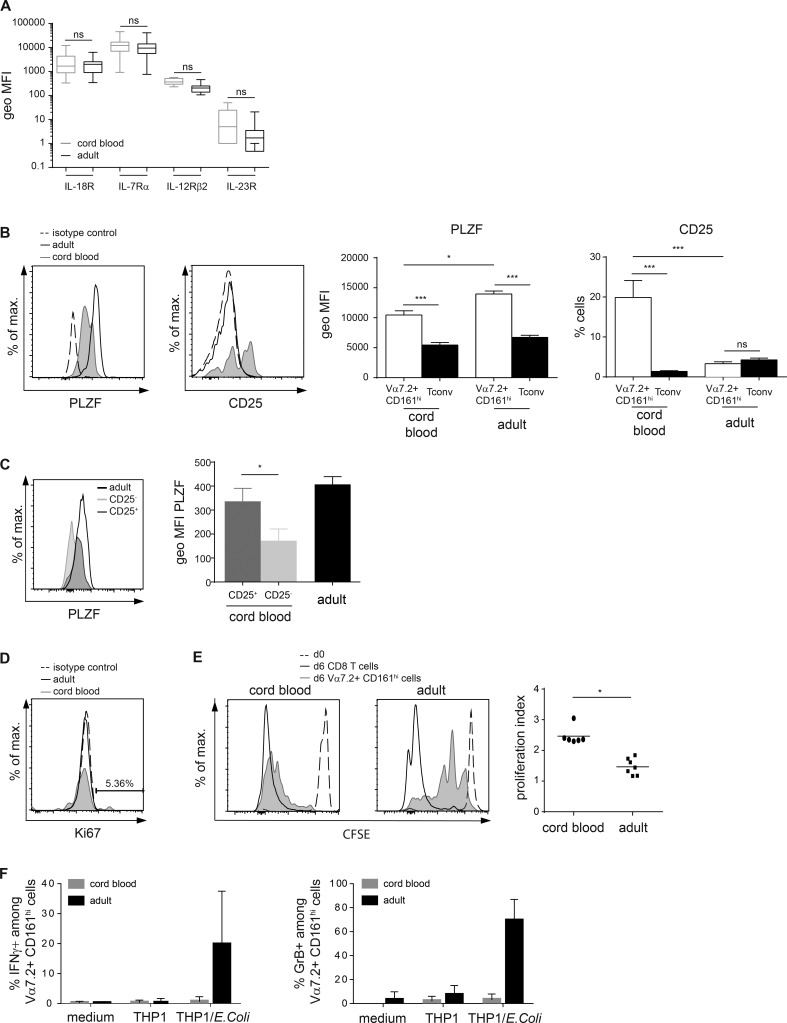
Mucosal-associated invariant T (MAIT) cells are semi-invariant Vα7.2+ CD161highCD4- T cells that recognize microbial riboflavin precursor derivatives such as 5-OP-RU presented by MR1. Human MAIT cells are abundant in adult blood, but there are very few in cord blood. We longitudinally studied Vα7.2+ CD161high T cell and related subset levels in infancy and after cord blood transplantation. We show that Vα7.2+ and Vα7.2- CD161high T cells are generated early during gestation and likely share a common prenatal developmental program. Among cord blood Vα7.2+ CD161high T cells, the minority recognizing MR1:5-OP-RU display a TRAV/TRBV repertoire very similar to adult MAIT cells. Within a few weeks of life, only the MR1:5-OP-RU reactive Vα7.2+ CD161high T cells acquire a memory phenotype. Only these cells expand to form the adult MAIT pool, diluting out other Vα7.2+ CD161high and Vα7.2- CD161high populations, in a process requiring at least 6 years to reach adult levels. Thus, the high clonal size of adult MAIT cells is antigen-driven and likely due to the fine specificity of the TCRαβ chains recognizing MR1-restricted microbial antigens.
© 2018 Ben Youssef et al.
Figures





References
-
- Abrahamsson S.V., Angelini D.F., Dubinsky A.N., Morel E., Oh U., Jones J.L., Carassiti D., Reynolds R., Salvetti M., Calabresi P.A., et al. 2013. Non-myeloablative autologous haematopoietic stem cell transplantation expands regulatory cells and depletes IL-17 producing mucosal-associated invariant T cells in multiple sclerosis. Brain. 136:2888–2903. 10.1093/brain/awt182 - DOI - PMC - PubMed
-
- Billerbeck E., Kang Y.H., Walker L., Lockstone H., Grafmueller S., Fleming V., Flint J., Willberg C.B., Bengsch B., Seigel B., et al. 2010. Analysis of CD161 expression on human CD8+ T cells defines a distinct functional subset with tissue-homing properties. Proc. Natl. Acad. Sci. USA. 107:3006–3011. 10.1073/pnas.0914839107 - DOI - PMC - PubMed
-
- Booth J.S., Salerno-Goncalves R., Blanchard T.G., Patil S.A., Kader H.A., Safta A.M., Morningstar L.M., Czinn S.J., Greenwald B.D., and Sztein M.B.. 2015. Mucosal-associated invariant T cells in the human gastric mucosa and blood: Role in helicobacter pylori infection. Front. Immunol. 6:466 10.3389/fimmu.2015.00466 - DOI - PMC - PubMed
-
- Chen Z., Wang H., D’Souza C., Sun S., Kostenko L., Eckle S.B., Meehan B.S., Jackson D.C., Strugnell R.A., Cao H., et al. 2017. Mucosal-associated invariant T-cell activation and accumulation after in vivo infection depends on microbial riboflavin synthesis and co-stimulatory signals. Mucosal Immunol. 10:58–68. 10.1038/mi.2016.39 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
