

C3G promotes a selective release of angiogenic factors from activated mouse platelets to regulate angiogenesis and tumor metastasis
- PMID: 29340032
- PMCID: PMC5762300
- DOI: 10.18632/oncotarget.22339
C3G promotes a selective release of angiogenic factors from activated mouse platelets to regulate angiogenesis and tumor metastasis
Abstract
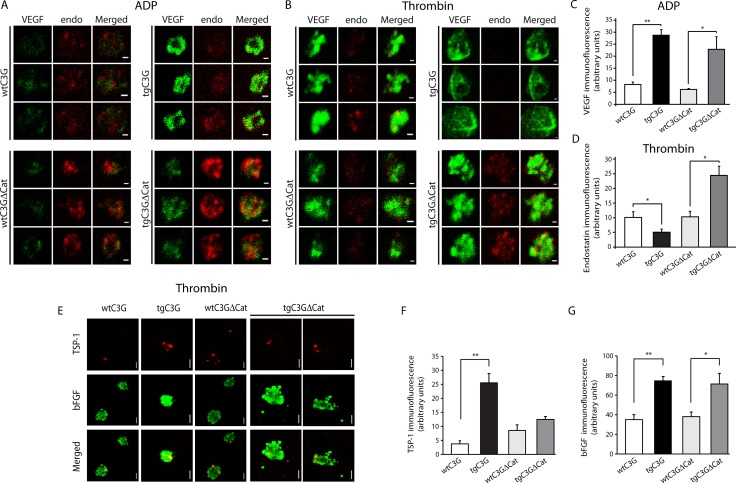
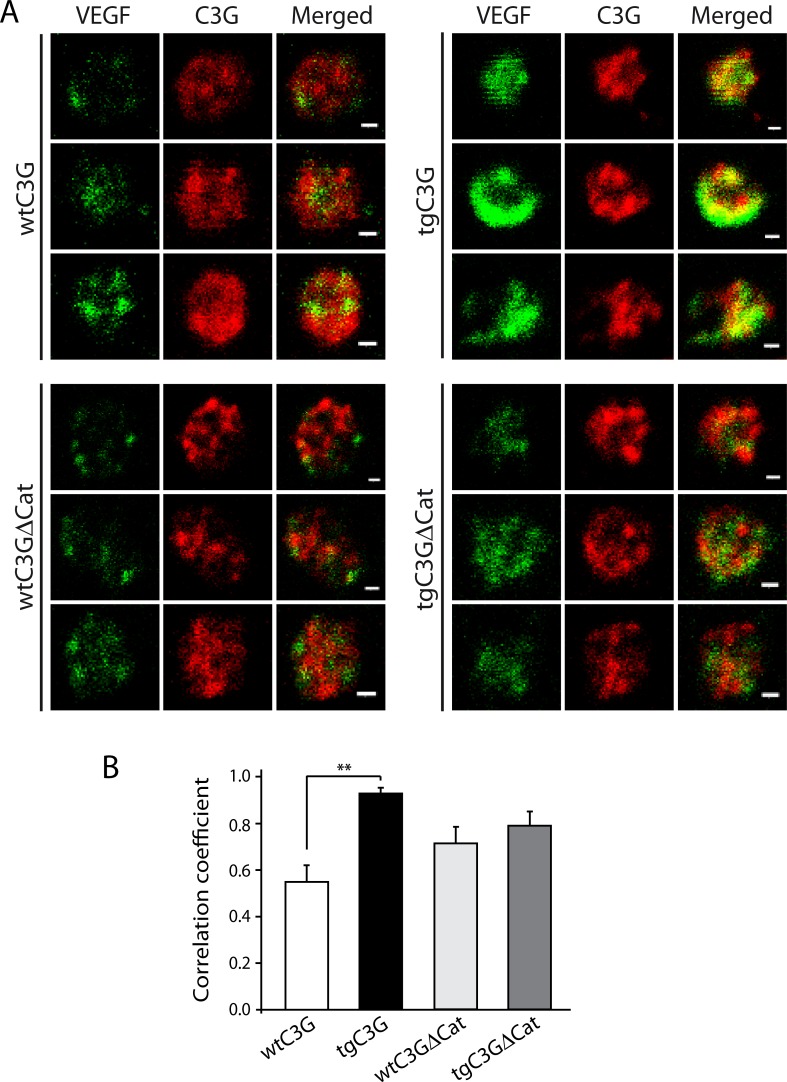
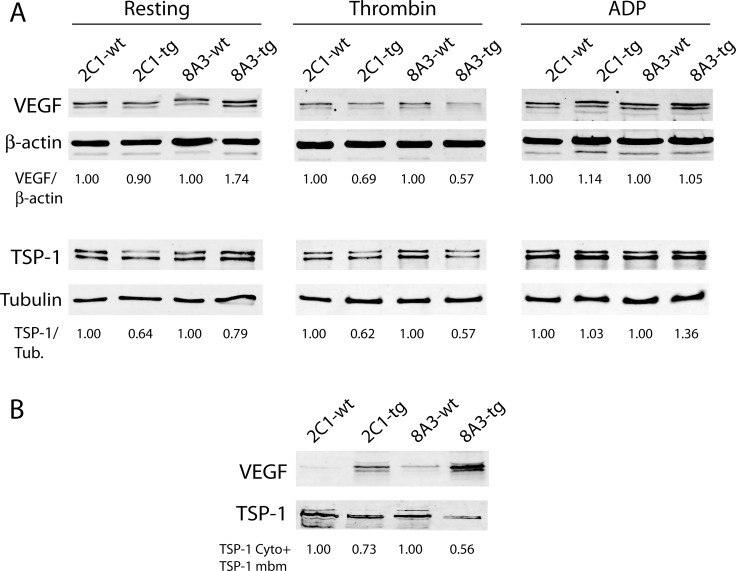
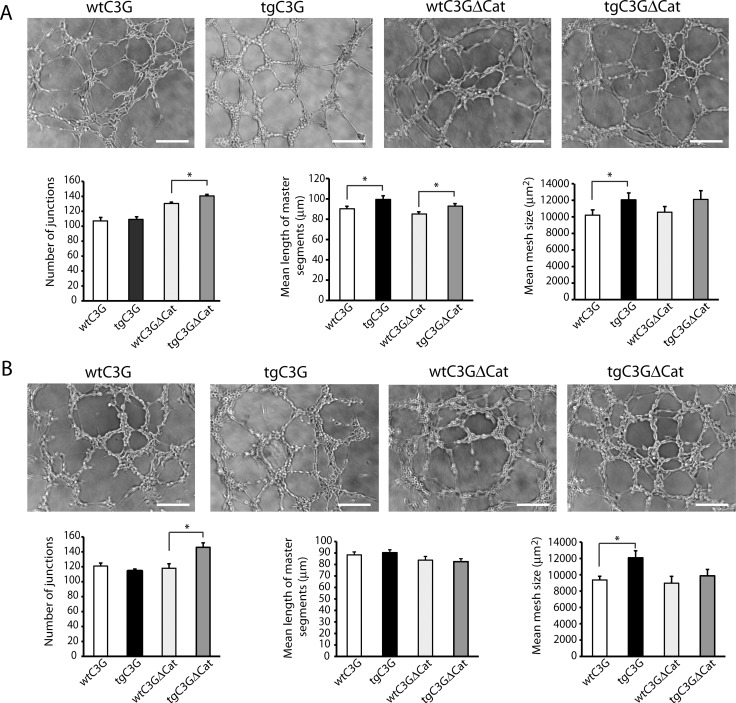
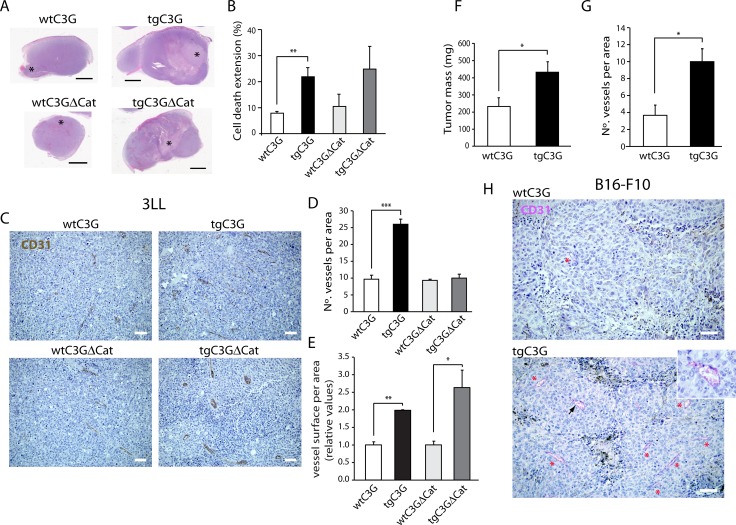
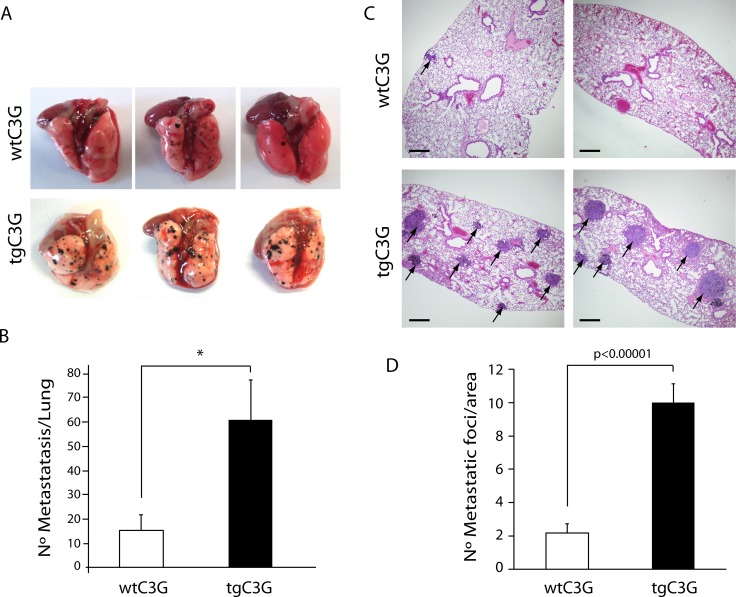
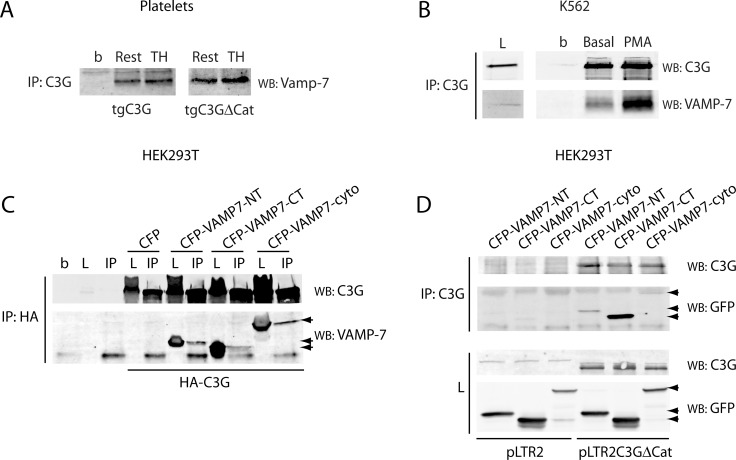
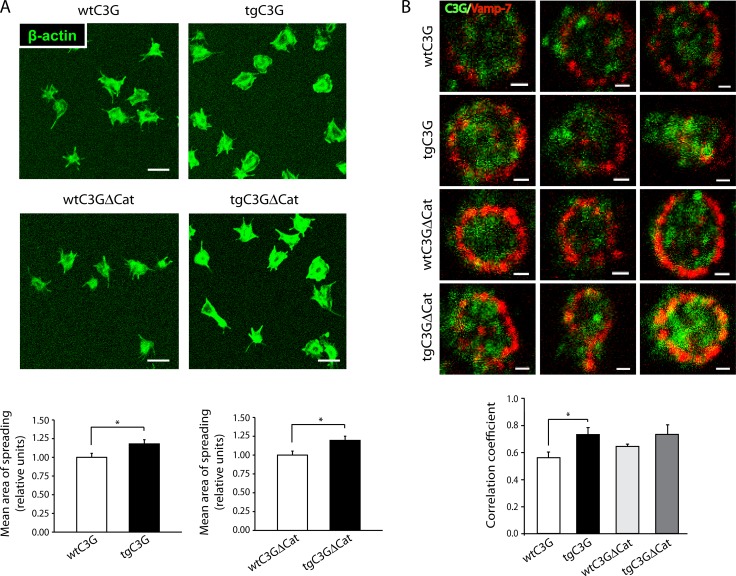
Previous observations indicated that C3G (RAPGEF1) promotes α-granule release, evidenced by the increase in P-selectin exposure on the platelet surface following its activation. The goal of the present study is to further characterize the potential function of C3G as a modulator of the platelet releasate and its implication in the regulation of angiogenesis. Proteomic analysis revealed a decreased secretion of anti-angiogenic factors from activated transgenic C3G and C3G∆Cat platelets. Accordingly, the secretome from both transgenic platelets had an overall pro-angiogenic effect as evidenced by an in vitro capillary-tube formation assay with HUVECs (human umbilical vein endothelial cells) and by two in vivo models of heterotopic tumor growth. In addition, transgenic C3G expression in platelets greatly increased mouse melanoma cells metastasis. Moreover, immunofluorescence microscopy showed that the pro-angiogenic factors VEGF and bFGF were partially retained into α-granules in thrombin- and ADP-activated mouse platelets from both, C3G and C3GΔCat transgenic mice. The observed interaction between C3G and Vesicle-associated membrane protein (Vamp)-7 could explain these results. Concomitantly, increased platelet spreading in both transgenic platelets upon thrombin activation supports this novel function of C3G in α-granule exocytosis. Collectively, our data point out to the co-existence of Rap1GEF-dependent and independent mechanisms mediating C3G effects on platelet secretion, which regulates pathological angiogenesis in tumors and other contexts. The results herein support an important role for platelet C3G in angiogenesis and metastasis.
Keywords: C3G; Vamp-7; angiogenesis; metastasis; platelet secretome.
Conflict of interest statement
CONFLICTS OF INTEREST The authors declare no competing financial interests.
Figures
References
-
- Guerrero C, Martin-Encabo S, Fernandez-Medarde A, Santos E. C3G-mediated suppression of oncogene-induced focus formation in fibroblasts involves inhibition of ERK activation, cyclin A expression and alterations of anchorage-independent growth. Oncogene. 2004;23:4885–4893. - PubMed
-
- Gutierrez-Berzal J, Castellano E, Martin-Encabo S, Gutierrez-Cianca N, Hernandez JM, Santos E, Guerrero C. Characterization of p87C3G, a novel, truncated C3G isoform that is overexpressed in chronic myeloid leukemia and interacts with Bcr-Abl. Exp Cell Res. 2006;312:938–948. - PubMed
-
- Martin-Encabo S, Santos E, Guerrero C. C3G mediated suppression of malignant transformation involves activation of PP2A phosphatases at the subcortical actin cytoskeleton. Exp Cell Res. 2007;313:3881–3891. - PubMed
-
- Maia V, Sanz M, Gutierrez-Berzal J, de Luis A, Gutierrez-Uzquiza A, Porras A, Guerrero C. C3G silencing enhances STI-571-induced apoptosis in CML cells through p38 MAPK activation, but it antagonizes STI-571 inhibitory effect on survival. Cell Signal. 2009;21:1229–1235. https://doi.org/10.1016/j.cellsig.2009.03.015. - DOI - PubMed
-
- Gutiérrez-Uzquiza A, Arechederra M, Molina I, Baños R, Maia V, Benito M, Guerrero C, Porras A. C3G down-regulates p38 MAPK activity in response to stress by Rap-1 independent mechanisms: Involvement in cell death. Cell Signal. 2010;22:533–542. https://doi.org/10.1016/j.cellsig.2009.11.008. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
