

Immunocontraceptive target repertoire defined by systematic identification of sperm membrane alloantigens in a single species
- PMID: 29342175
- PMCID: PMC5771590
- DOI: 10.1371/journal.pone.0190891
Immunocontraceptive target repertoire defined by systematic identification of sperm membrane alloantigens in a single species
Abstract
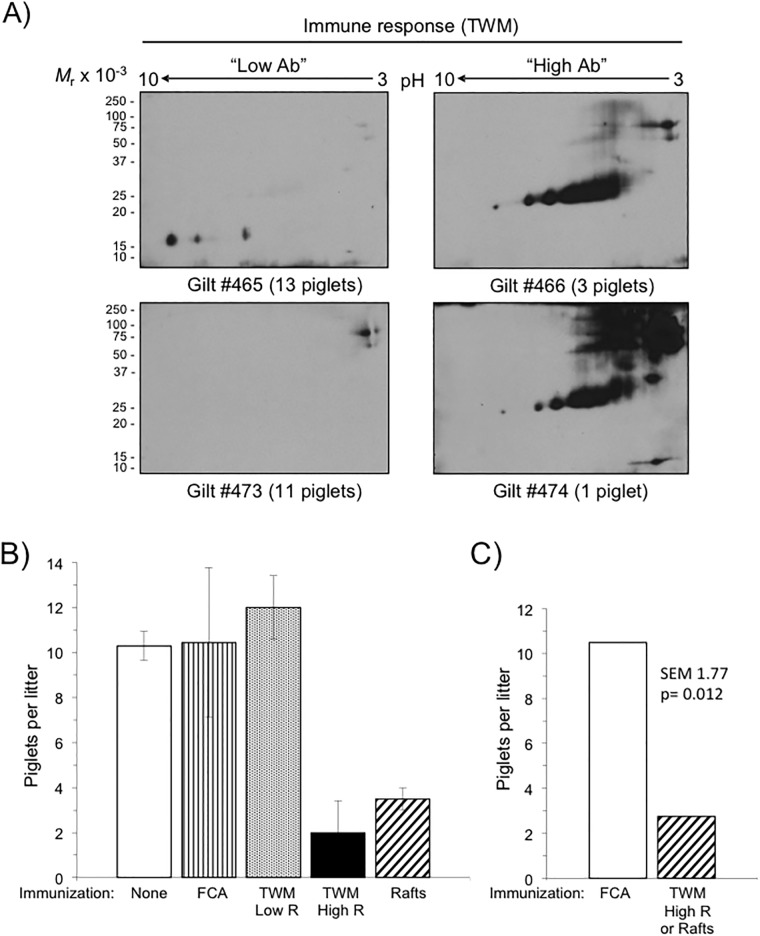
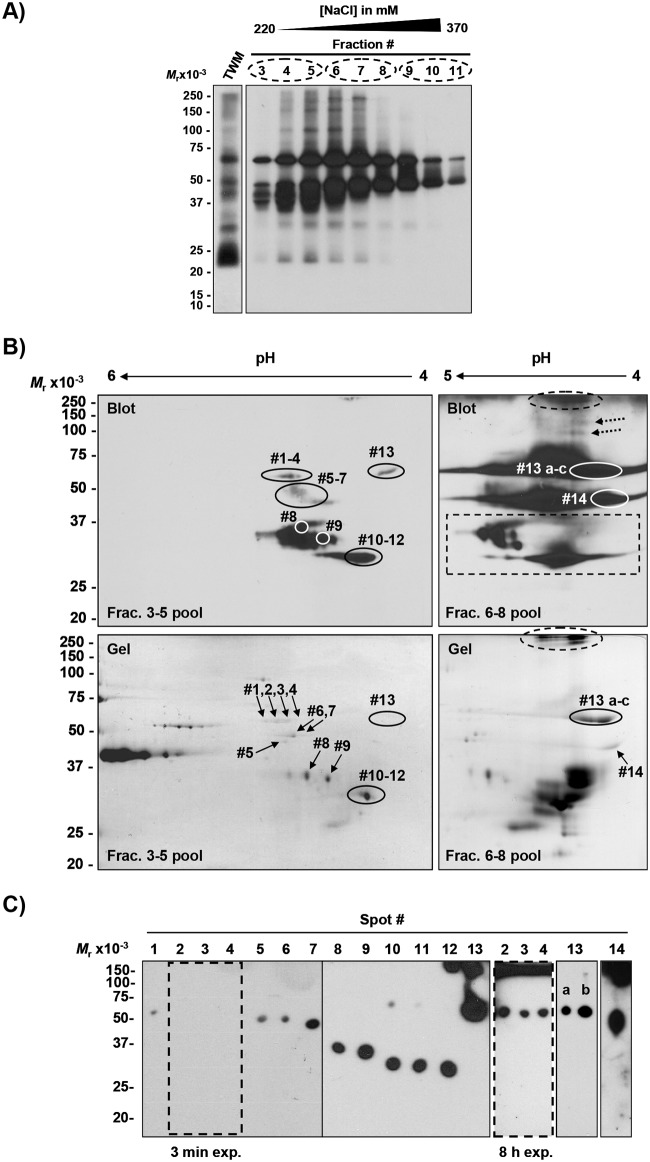
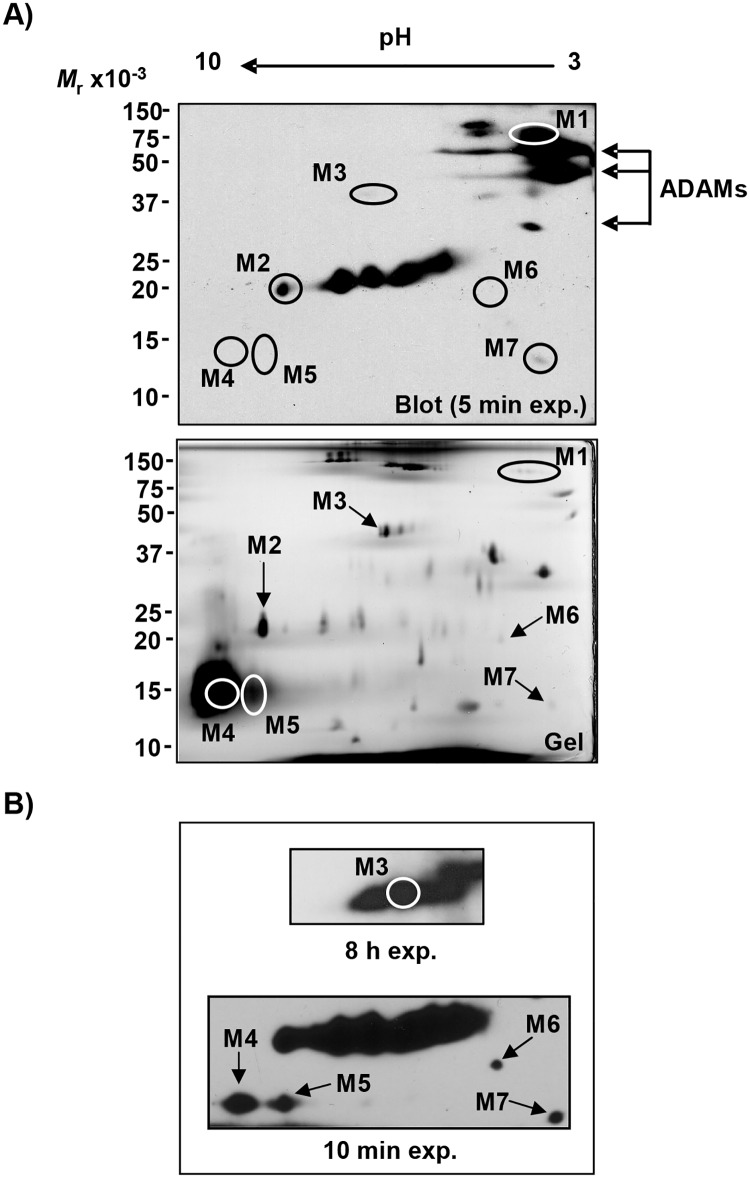
Sperm competence in animal fertilization requires the collective activities of numerous sperm-specific proteins that are typically alloimmunogenic in females. Consequently, sperm membrane alloantigens are potential targets for contraceptives that act by blocking the proteins' functions in gamete interactions. Here we used a targeted proteomics approach to identify the major alloantigens in swine sperm membranes and lipid rafts, and thereby systematically defined the repertoire of these sperm-specific proteins in a single species. Gilts with high alloantibody reactivity to proteins in sperm membranes or lipid rafts produced fewer offspring (73% decrease) than adjuvant-only or nonimmune control animals. Alloantisera recognized more than 20 potentially unique sperm membrane proteins and five sperm lipid raft proteins resolved on two-dimensional immunoblots with or without prior enrichment by anion exchange chromatography. Dominant sperm membrane alloantigens identified by mass spectrometry included the ADAMs fertilin α, fertilin ß, and cyritestin. Less abundant alloantigens included ATP synthase F1 β subunit, myo-inositol monophosphatase-1, and zymogen granule membrane glycoprotein-2. Immunodominant sperm lipid raft alloantigens included SAMP14, lymphocyte antigen 6K, and the epididymal sperm protein E12. Of the fifteen unique membrane alloantigens identified, eleven were known sperm-specific proteins with uncertain functions in fertilization, and four were not previously suspected to exist as sperm-specific isoforms. De novo sequences of tryptic peptides from sperm membrane alloantigen "M6" displayed no evident homology to known proteins, so is a newly discovered sperm-specific gene product in swine. We conclude that alloimmunizing gilts with sperm membranes or lipid rafts evokes formation of antibodies to a relatively small number of dominant alloantigens that include known and novel sperm-specific proteins with possible functions in fertilization and potential utility as targets for immunocontraception.
Conflict of interest statement
Figures


Similar articles
-
Systematic characterization of sperm-specific membrane proteins in swine.Biol Reprod. 2000 Dec;63(6):1839-47. doi: 10.1095/biolreprod63.6.1839. Biol Reprod. 2000. PMID: 11090456
-
Capacitation-dependent concentration of lipid rafts in the apical ridge head area of porcine sperm cells.Mol Hum Reprod. 2005 Aug;11(8):583-90. doi: 10.1093/molehr/gah200. Epub 2005 Jul 28. Mol Hum Reprod. 2005. PMID: 16051681
-
Examination of the immunocontraceptive potential of recombinant rabbit fertilin subunits in rabbit.Biol Reprod. 1997 Oct;57(4):879-86. doi: 10.1095/biolreprod57.4.879. Biol Reprod. 1997. PMID: 9314593
-
Sperm antigens and their use in the development of an immunocontraceptive.Am J Reprod Immunol. 1997 Jan;37(1):111-7. doi: 10.1111/j.1600-0897.1997.tb00199.x. Am J Reprod Immunol. 1997. PMID: 9138444 Review.
-
Identification of sperm immunoreactive antigens for immunocontraceptive purposes: a review.Reprod Biol Endocrinol. 2004 Mar 18;2:11. doi: 10.1186/1477-7827-2-11. Reprod Biol Endocrinol. 2004. PMID: 15035665 Free PMC article. Review.
Cited by
-
The novel BRDT inhibitor NHWD870 shows potential as a male contraceptive in mice.Acta Biochim Biophys Sin (Shanghai). 2022 Sep 25;54(12):1789-1800. doi: 10.3724/abbs.2022135. Acta Biochim Biophys Sin (Shanghai). 2022. PMID: 36239350 Free PMC article.
-
SMA20/PMIS2 Is a Rapidly Evolving Sperm Membrane Alloantigen with Possible Species-Divergent Function in Fertilization.Int J Mol Sci. 2024 Mar 25;25(7):3652. doi: 10.3390/ijms25073652. Int J Mol Sci. 2024. PMID: 38612464 Free PMC article.
-
Update on Novel Hormonal and Nonhormonal Male Contraceptive Development.J Clin Endocrinol Metab. 2021 May 13;106(6):e2381-e2392. doi: 10.1210/clinem/dgab034. J Clin Endocrinol Metab. 2021. PMID: 33481994 Free PMC article. Review.
References
-
- Aitken RJ.Immunocontraceptive vaccines for human use. J Reprod Immunol. 2002;57: 273–287. - PubMed
-
- Bradley MP, Bird P Applications of fertility regulation for the management of wild and domestic species In: Hardy DM, editor. Fertilization. Academic Press, San Diego; 2002. pp. 401–418.
-
- Haden NP, Hickox JR, Whisnant CS, Hardy DM Systematic characterization of sperm-specific membrane proteins in swine. Biol Reprod. 2000;63: 1839–1847. - PubMed
-
- Druce HC, Mackey RL, Slotow R How immunocontraception can contribute to elephant management in small, enclosed reserves: Munyawana population as a case study. PLoS One. 2011;6: e27952 doi: 10.1371/journal.pone.0027952 - DOI - PMC - PubMed
-
- Goldberg E. Infertility in female rabbits immunized with lactate dehydrogenase X. Science. 1973;181: 458–459. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
