

Canonical and Noncanonical Actions of Arabidopsis Histone Deacetylases in Ribosomal RNA Processing
- PMID: 29343504
- PMCID: PMC5810568
- DOI: 10.1105/tpc.17.00626
Canonical and Noncanonical Actions of Arabidopsis Histone Deacetylases in Ribosomal RNA Processing
Abstract
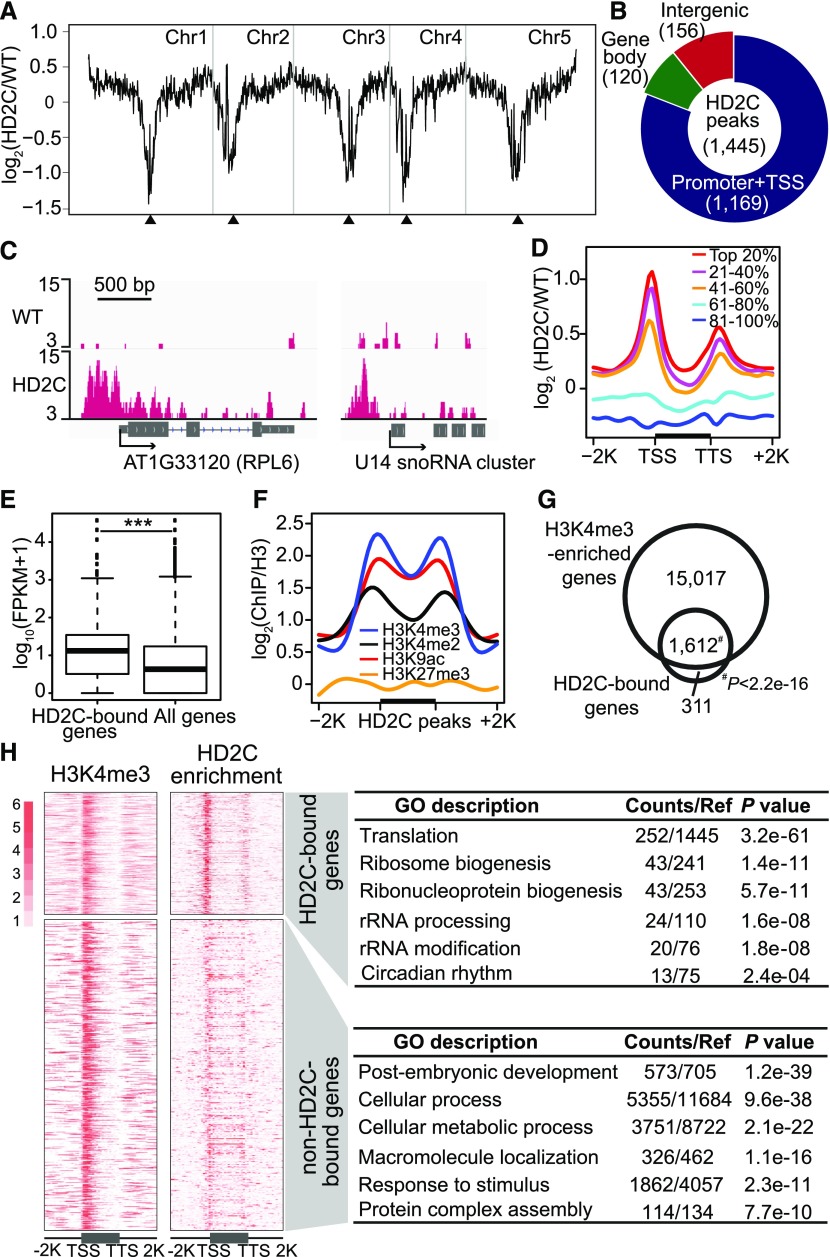
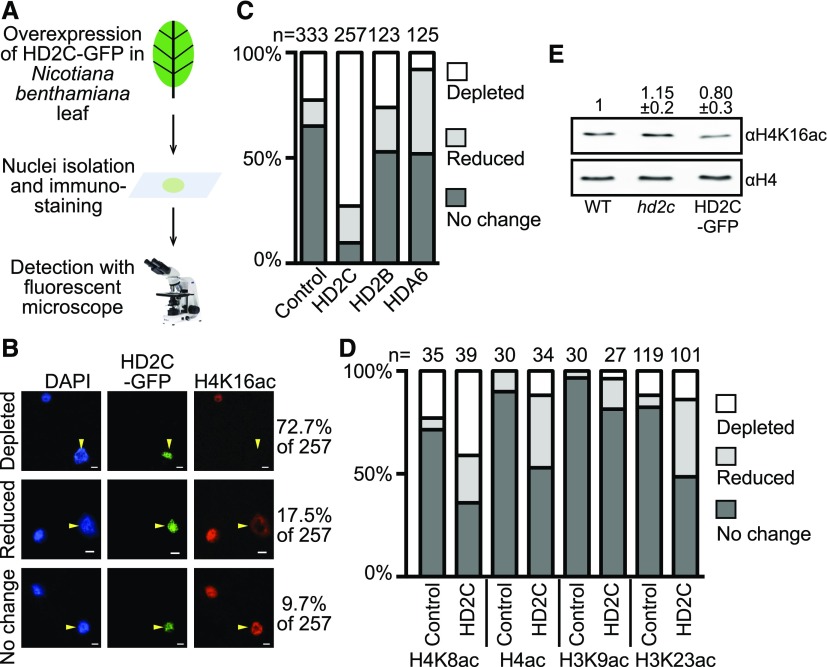
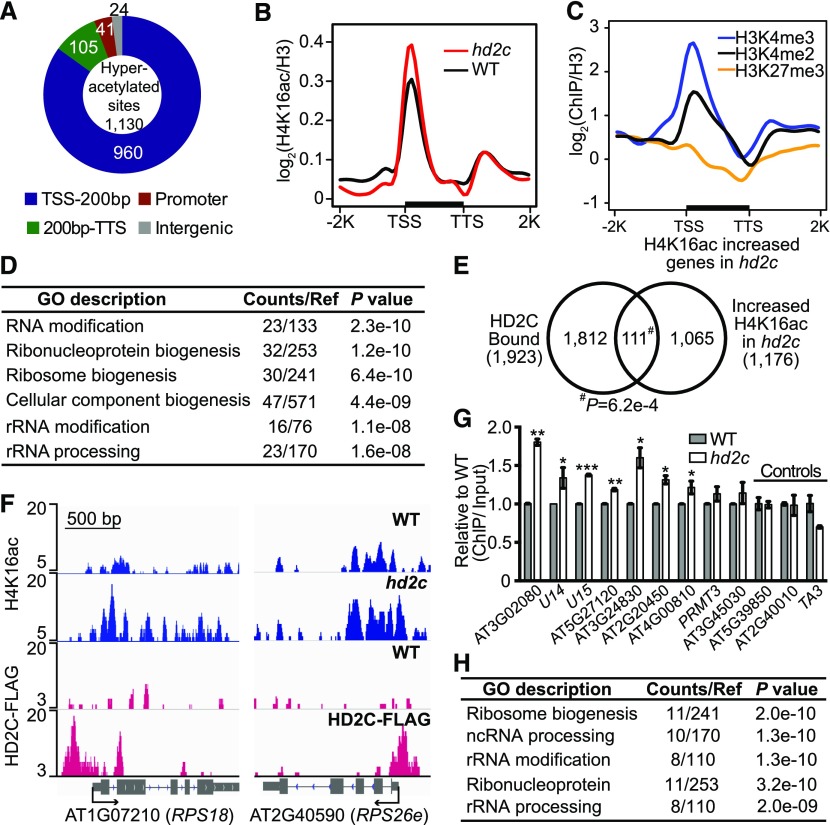
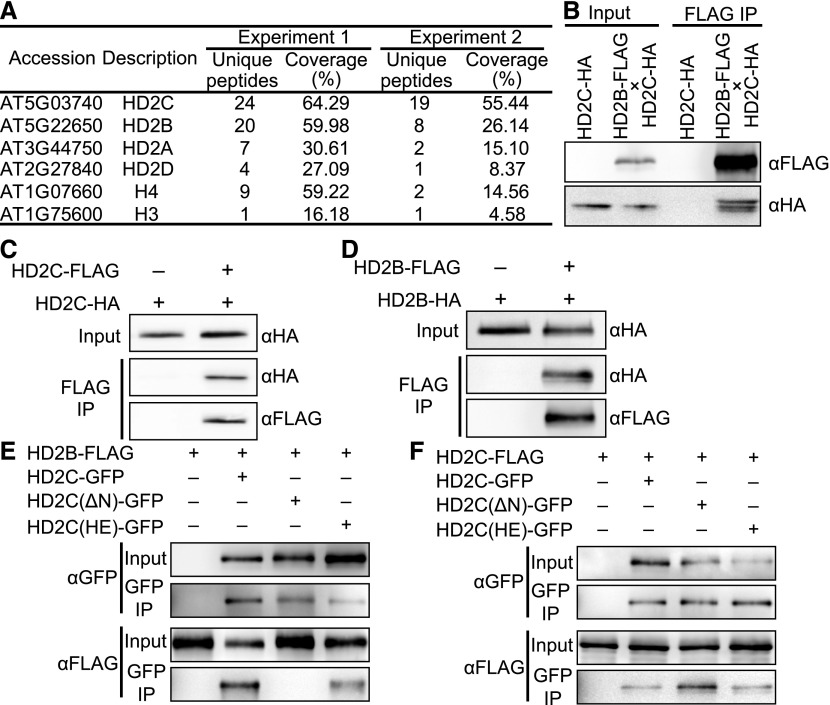
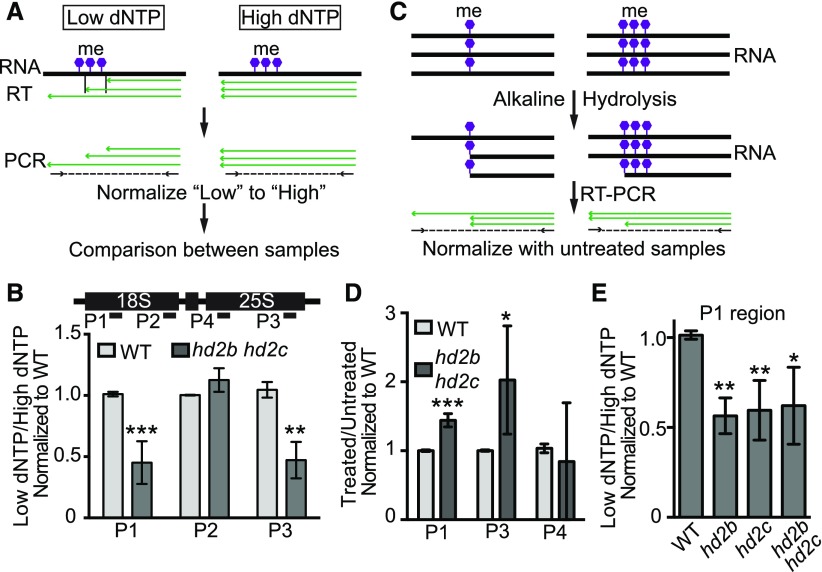
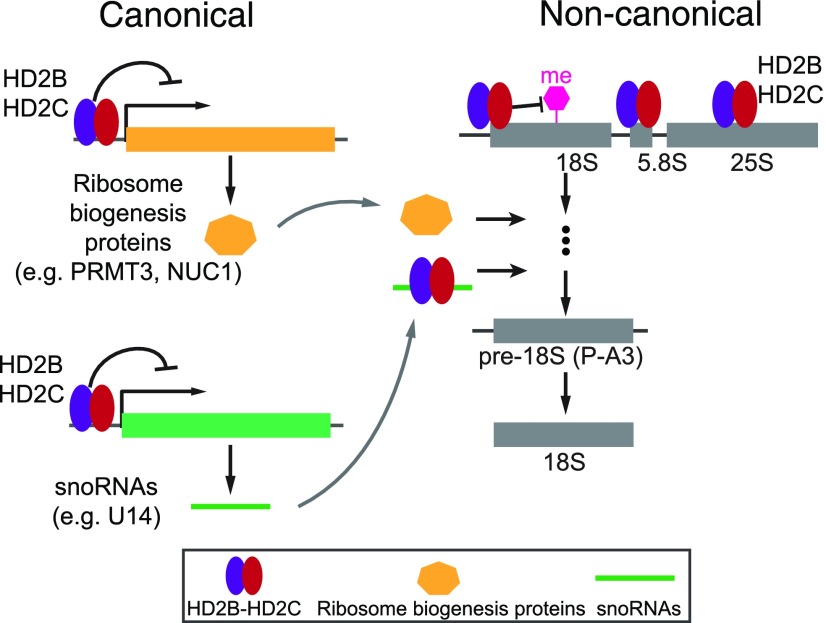
Ribosome biogenesis is a fundamental process required for all cellular activities. Histone deacetylases play critical roles in many biological processes including transcriptional repression and rDNA silencing. However, their function in pre-rRNA processing remains poorly understood. Here, we discovered a previously uncharacterized role of Arabidopsis thaliana histone deacetylase HD2C in pre-rRNA processing via both canonical and noncanonical manners. HD2C interacts with another histone deacetylase HD2B and forms homo- and/or hetero-oligomers in the nucleolus. Depletion of HD2C and HD2B induces a ribosome-biogenesis deficient phenotype and aberrant accumulation of 18S pre-rRNA intermediates. Our genome-wide analysis revealed that HD2C binds and represses the expression of key genes involved in ribosome biogenesis. Using RNA immunoprecipitation and sequencing, we further uncovered a noncanonical mechanism of HD2C directly associating with pre-rRNA and small nucleolar RNAs to regulate rRNA methylation. Together, this study reveals a multifaceted role of HD2C in ribosome biogenesis and provides mechanistic insights into how histone deacetylases modulate rRNA maturation at the transcriptional and posttranscriptional levels.
© 2018 American Society of Plant Biologists. All rights reserved.
Figures
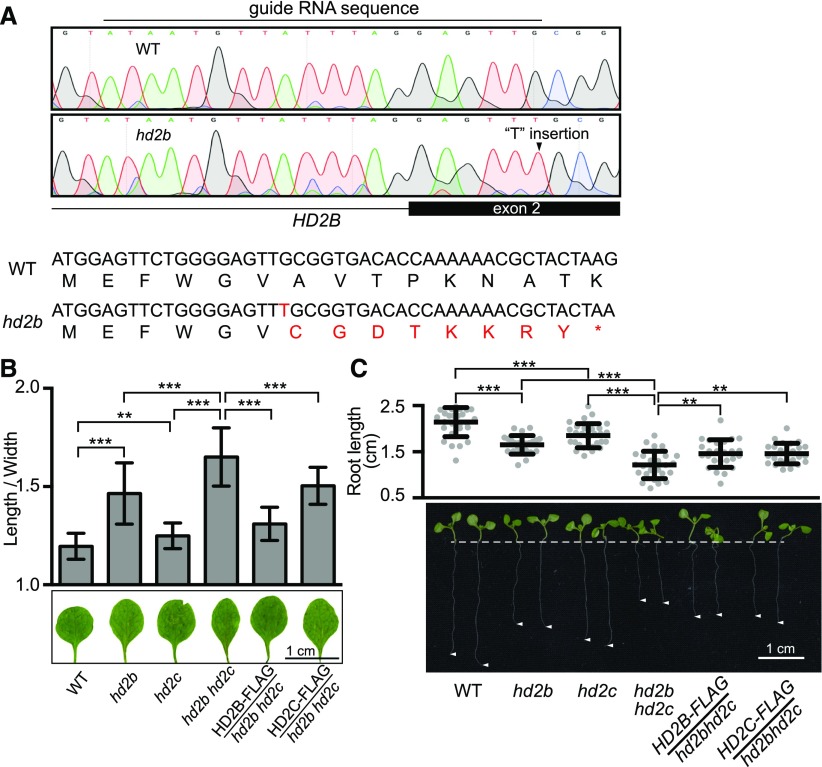
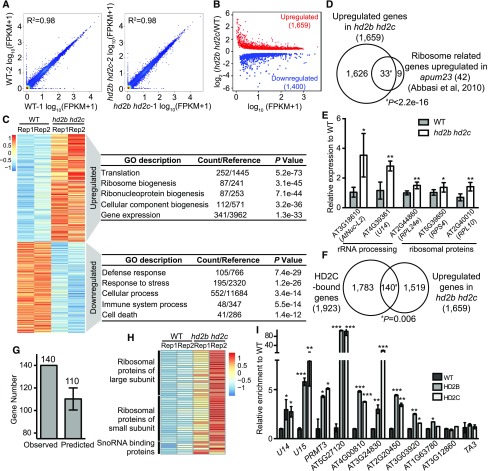
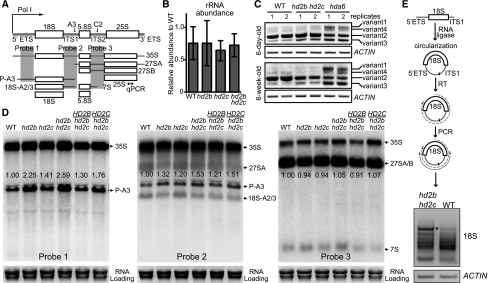
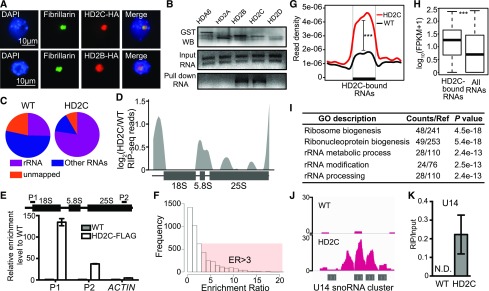
References
-
- Abbasi N., Kim H.B., Park N.I., Kim H.S., Kim Y.K., Park Y.I., Choi S.B. (2010). APUM23, a nucleolar Puf domain protein, is involved in pre-ribosomal RNA processing and normal growth patterning in Arabidopsis. Plant J. 64: 960–976. - PubMed
-
- Anantharaman V., Koonin E.V., Aravind L. (2002). SPOUT: a class of methyltransferases that includes spoU and trmD RNA methylase superfamilies, and novel superfamilies of predicted prokaryotic RNA methylases. J. Mol. Microbiol. Biotechnol. 4: 71–75. - PubMed
-
- Aravind L., Koonin E.V. (1998). Second family of histone deacetylases. Science 280: 1167a.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
