

Frontline Science: High fat diet and leptin promote tumor progression by inducing myeloid-derived suppressor cells
- PMID: 29345342
- PMCID: PMC7414791
- DOI: 10.1002/JLB.4HI0517-210R
Frontline Science: High fat diet and leptin promote tumor progression by inducing myeloid-derived suppressor cells
Abstract
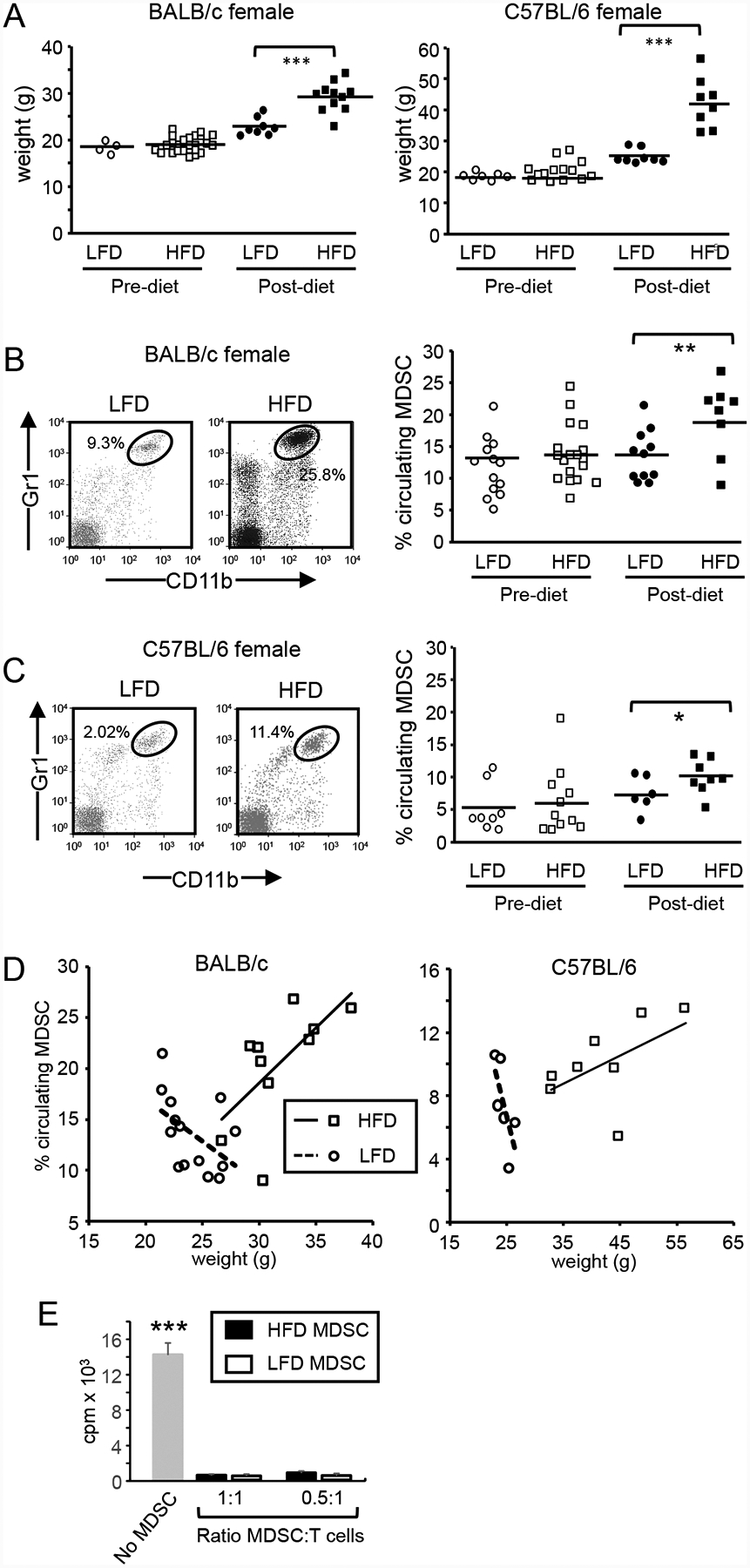
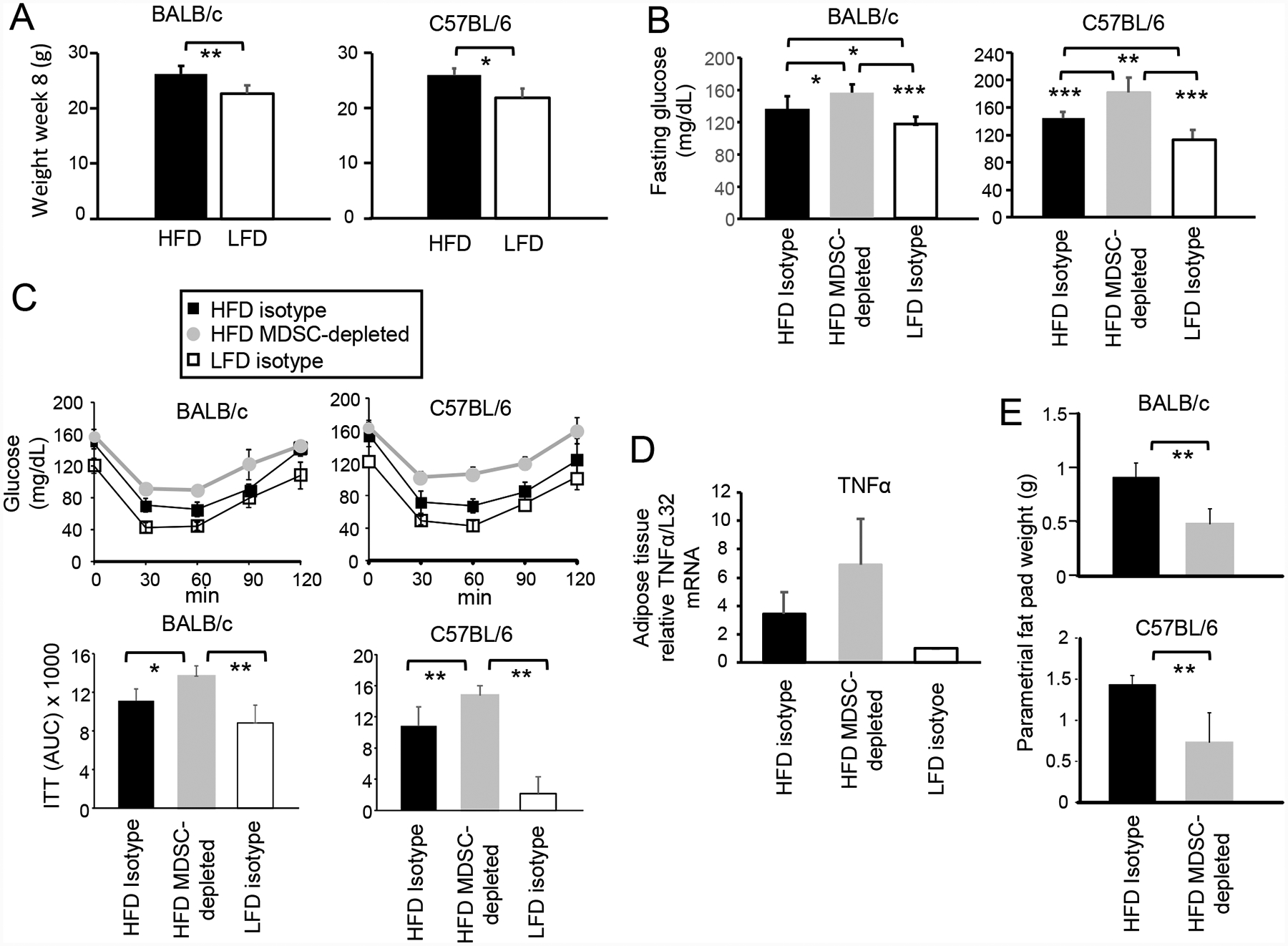
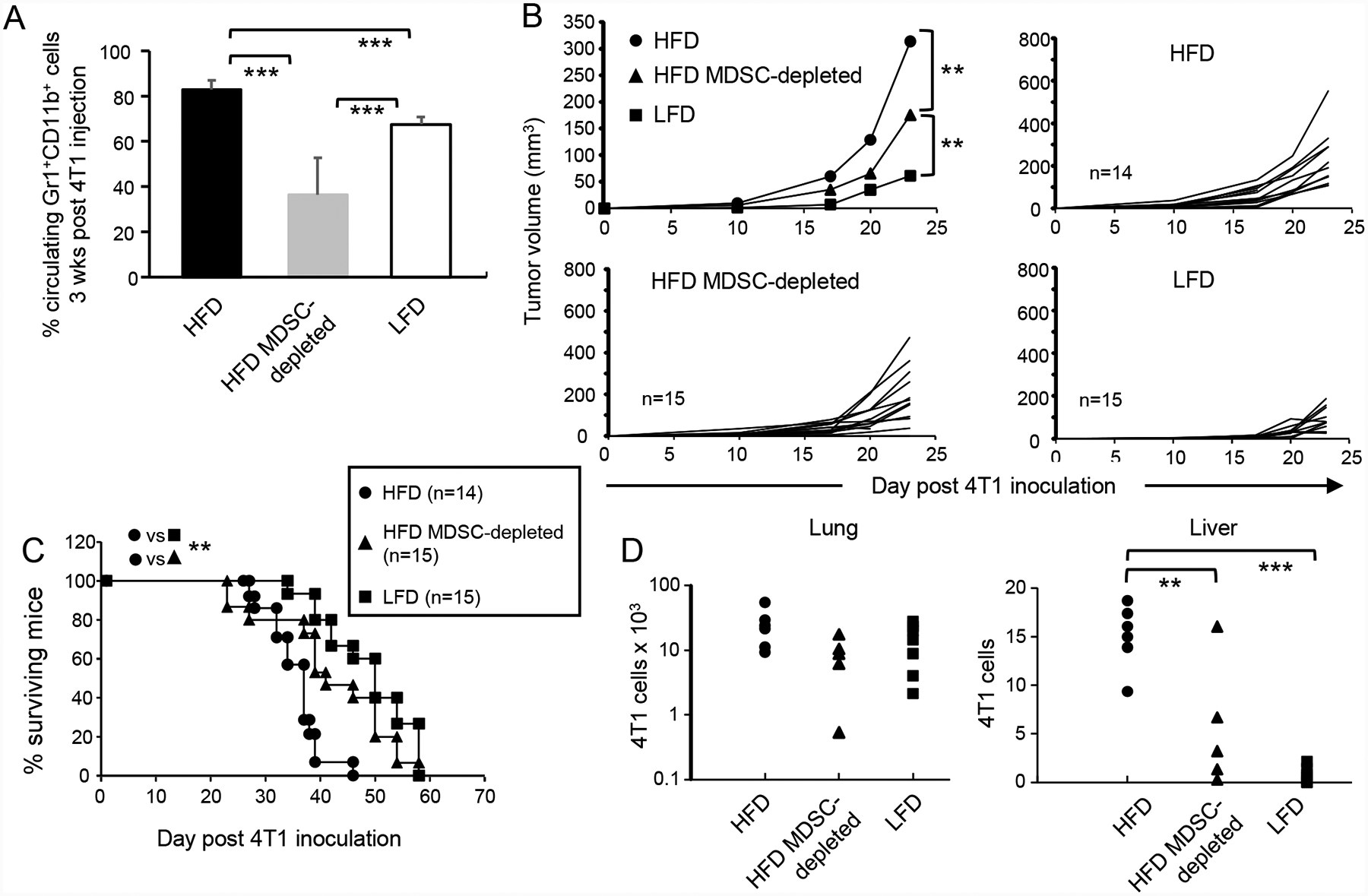
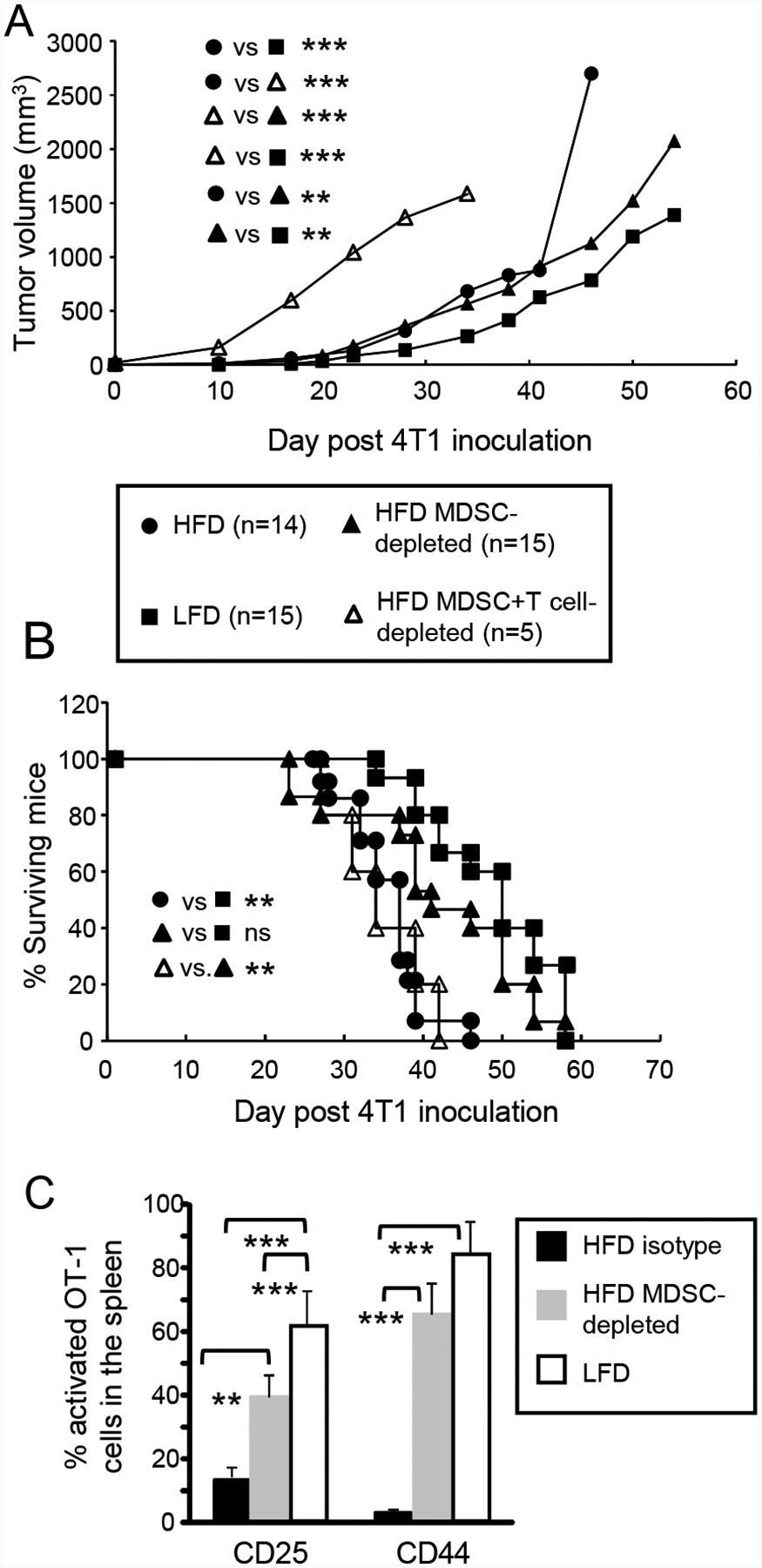
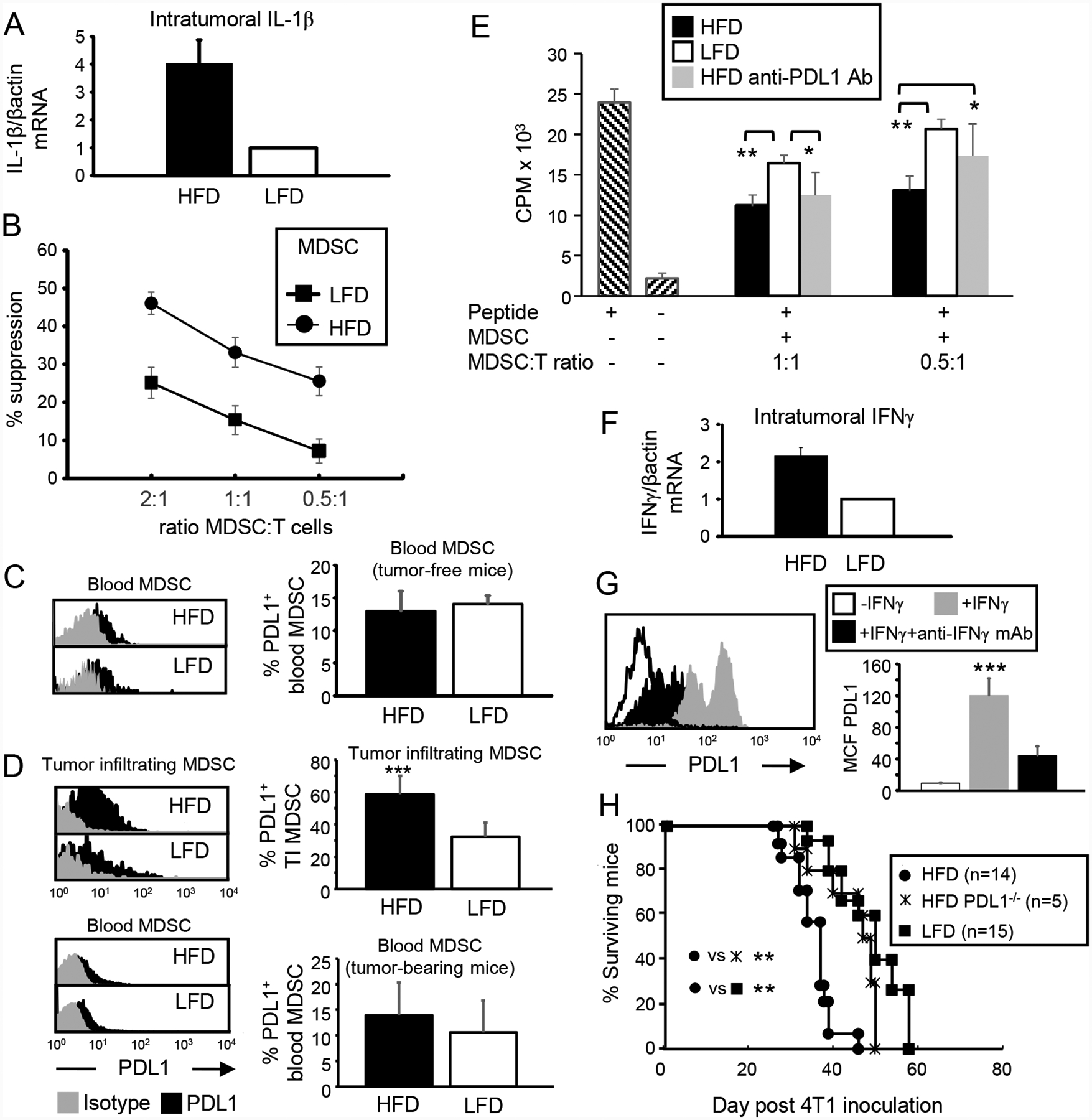
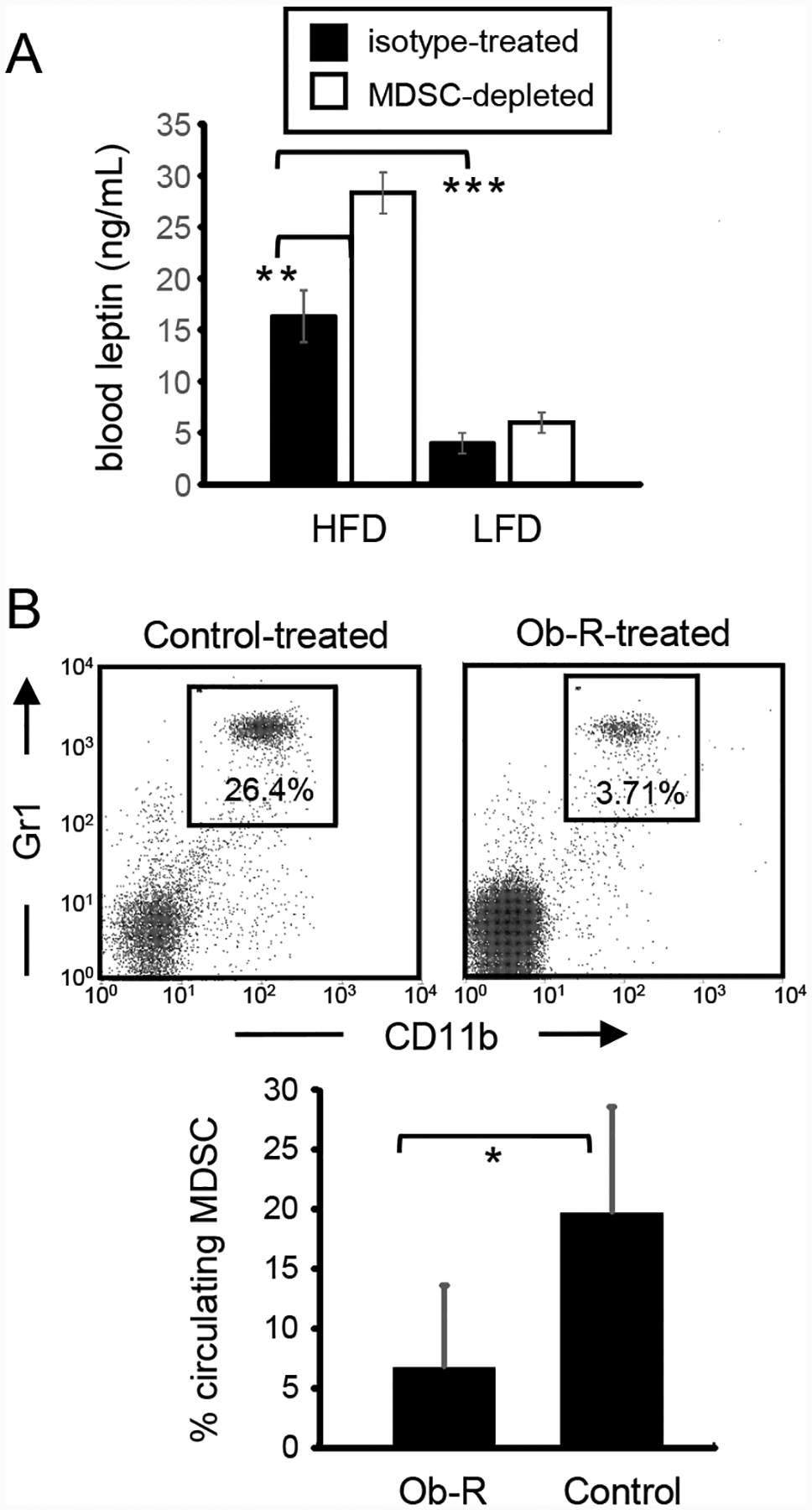
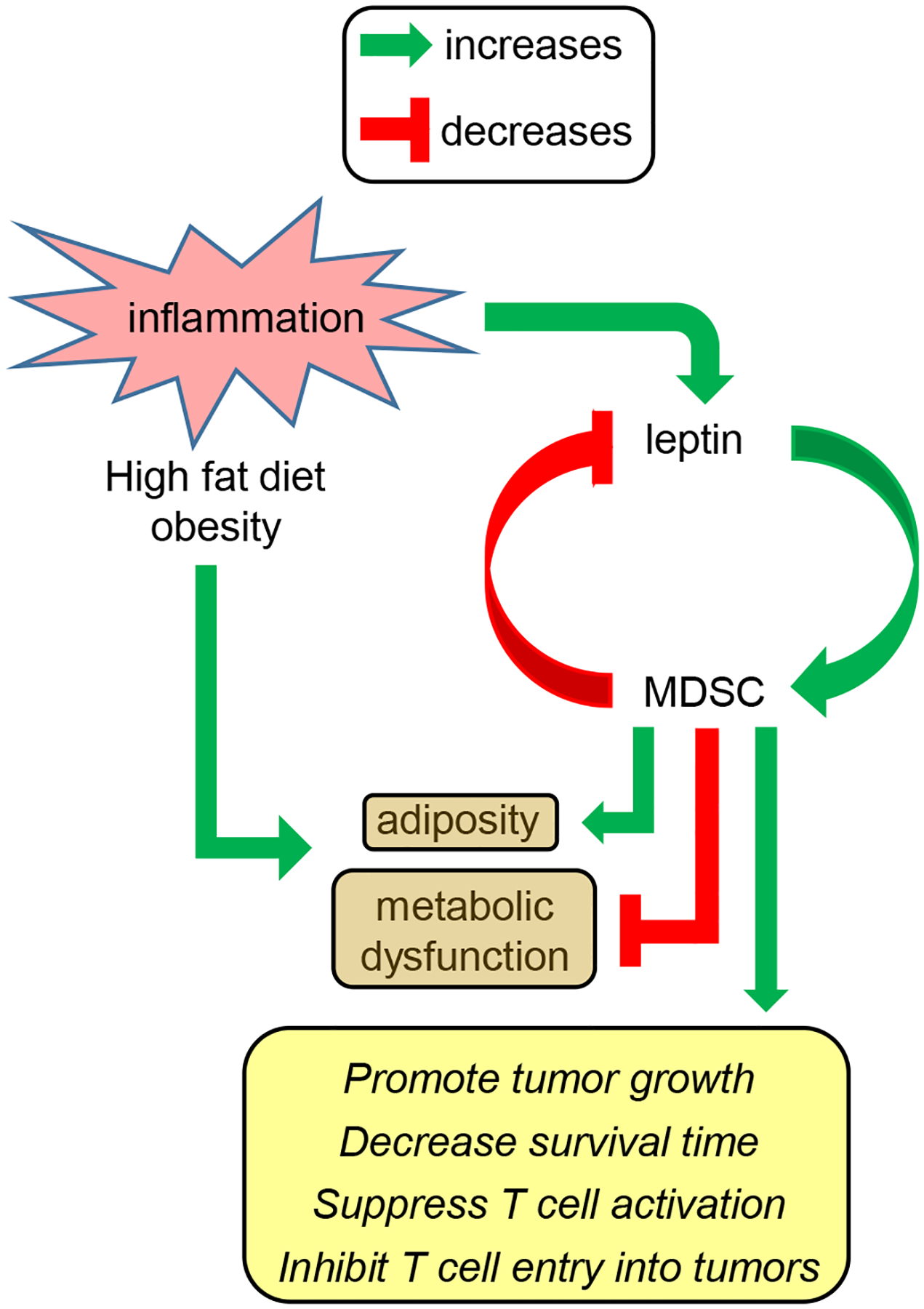
Obesity is a risk factor for cancer incidence and cancer mortality. The association of obesity and cancer is attributed to multiple factors, but the tightest linkage is with the chronic, low-grade inflammation that accompanies obesity. Myeloid-derived suppressor cells (MDSC) are known facilitators of cancer progression that act by suppressing the activation and function of tumor-reactive T cells. Because MDSC quantity and function are driven by chronic inflammation, we hypothesized that MDSC may accumulate in obese individuals and facilitate tumor growth by suppressing antitumor immunity. To test this hypothesis, tumor-bearing mice on a high fat or low fat diet (HFD or LFD) were assessed for tumor progression and the metabolic dysfunction associated with obesity. HFD enhanced the accumulation of MDSC, and the resulting MDSC had both beneficial and detrimental effects. HFD-induced MDSC protected mice against diet-induced metabolic dysfunction and reduced HFD-associated inflammation, but also increased the accumulation of fat, enhanced tumor progression, and spontaneous metastasis and reduced survival time. HFD-induced MDSC facilitated tumor growth by limiting the activation of tumor-reactive CD8+ T cells. Leptin, an adipokine that regulates appetite satiety and is overexpressed in obesity, undergoes crosstalk with MDSC in which leptin drives the accumulation of MDSC while MDSC down-regulate the production of leptin. Collectively, these studies demonstrate that although MDSC protect against some metabolic dysfunction associated with HFD they enhance tumor growth in HFD mice and that leptin is a key regulator linking HFD, chronic inflammation, immune suppression, and tumor progression.
Keywords: immune suppression; inflammation; obesity; programmed death ligand 1.
©2018 Society for Leukocyte Biology.
Conflict of interest statement
Conflict of Interest Disclosure
The authors declare no conflicts of interest
Figures
Comment in
-
Editorial: Two MDSC faces in obesity: Correcting metabolic dysfunctions but promoting tumor development.J Leukoc Biol. 2018 Mar;103(3):373-375. doi: 10.1002/JLB.3CE1017-416R. Epub 2017 Dec 15. J Leukoc Biol. 2018. PMID: 29345358 No abstract available.
References
-
- Calle EE, Rodriguez C, Walker-Thurmond K, Thun MJ (2003) Overweight, obesity, and mortality from cancer in a prospectively studied cohort of U.S. adults. N. Engl. J. Med 348, 1625–38. - PubMed
-
- Renehan AG, Tyson M, Egger M, Heller RF, Zwahlen M (2008) Body-mass index and incidence of cancer: a systematic review and meta-analysis of prospective observational studies. Lancet 371, 569–78. - PubMed
-
- Font-Burgada J, Sun B, Karin M (2016) Obesity and Cancer: The Oil that Feeds the Flame. Cell. Metab 23, 48–62. - PubMed
-
- van Kruijsdijk RC, van der Wall E, Visseren FL (2009) Obesity and cancer: the role of dysfunctional adipose tissue. Cancer Epidemiol. Biomarkers Prev 18, 2569–78. - PubMed
-
- Feola A, Ricci S, Kouidhi S, Rizzo A, Penon A, Formisano P, Giordano A, Di Carlo A, Di Domenico M (2017) Multifaceted Breast Cancer: The Molecular Connection With Obesity. J. Cell. Physiol 232, 69–77. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
