

Multimodal Encoding of Novelty, Reward, and Learning in the Primate Nucleus Basalis of Meynert
- PMID: 29348191
- PMCID: PMC5824738
- DOI: 10.1523/JNEUROSCI.2021-17.2017
Multimodal Encoding of Novelty, Reward, and Learning in the Primate Nucleus Basalis of Meynert
Abstract
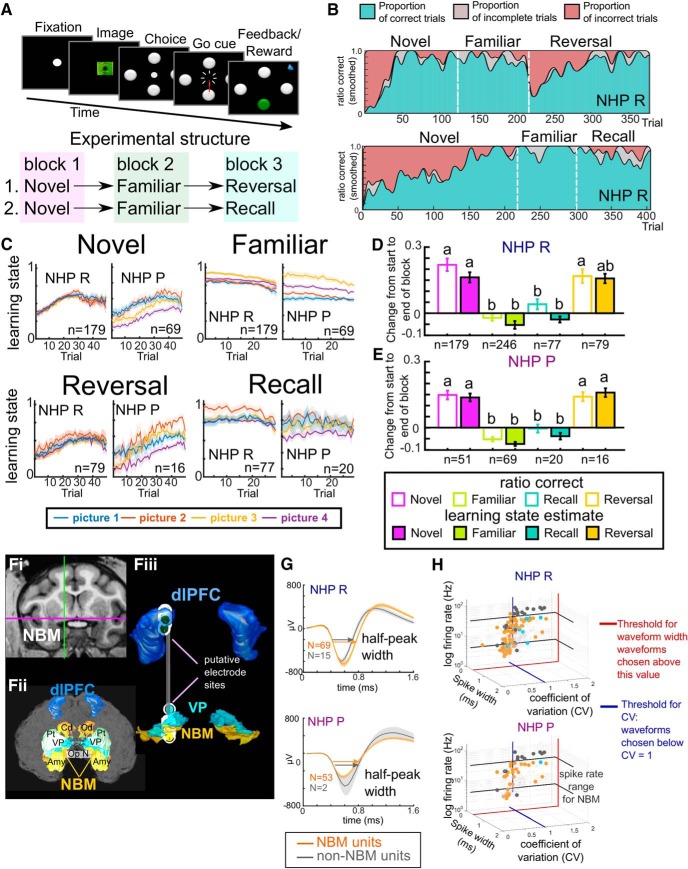
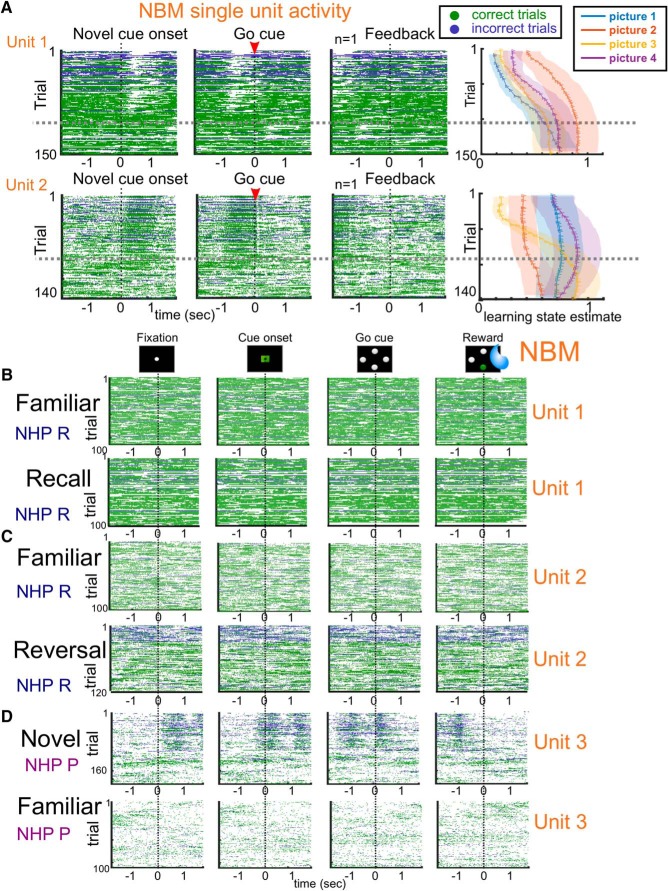
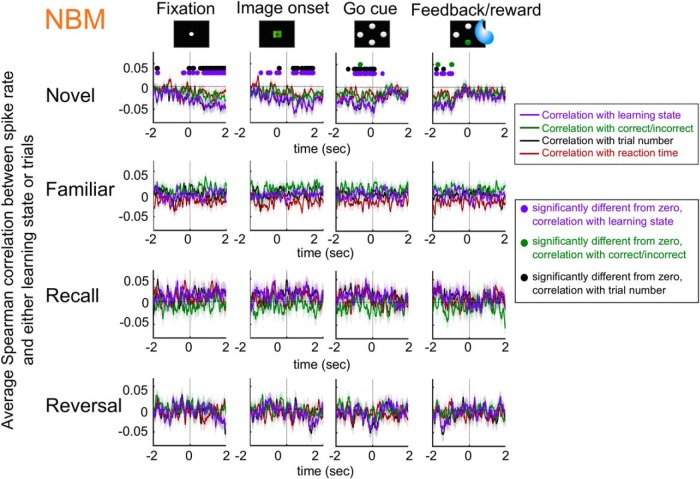
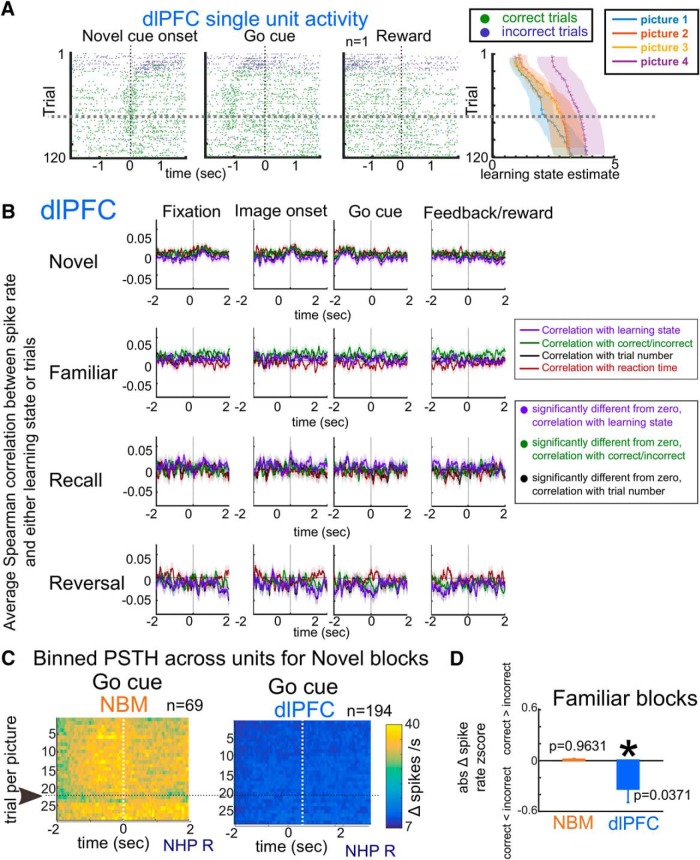
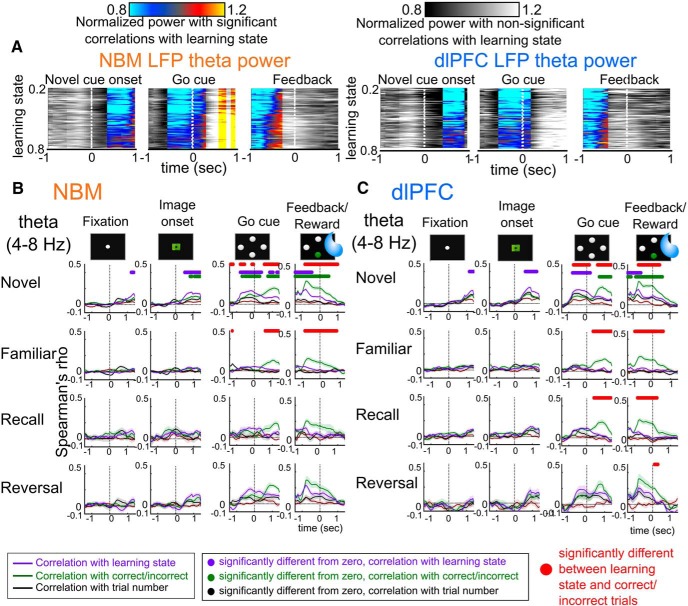
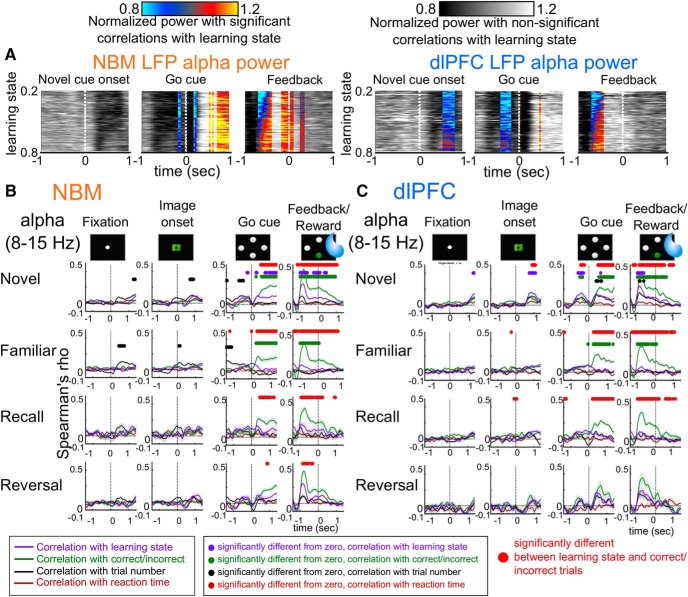
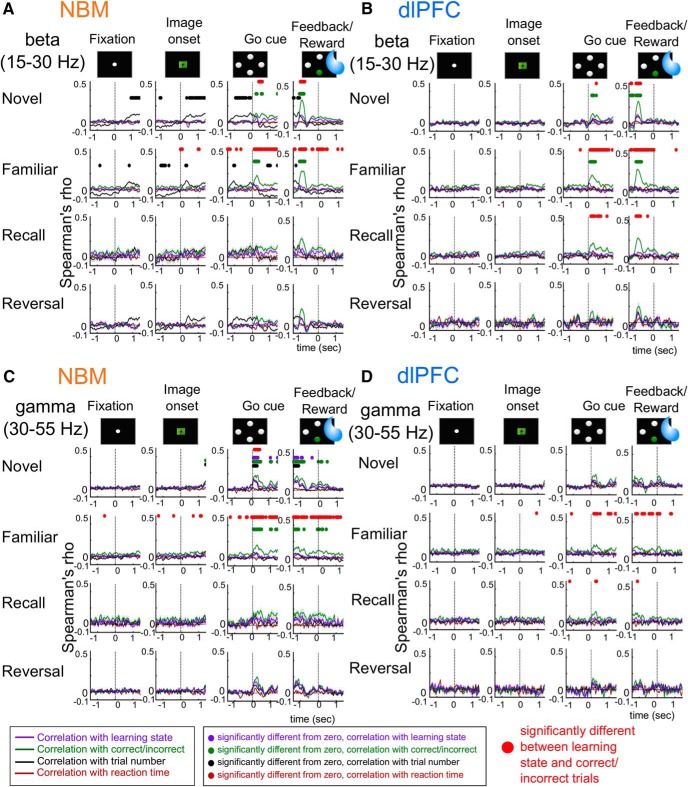
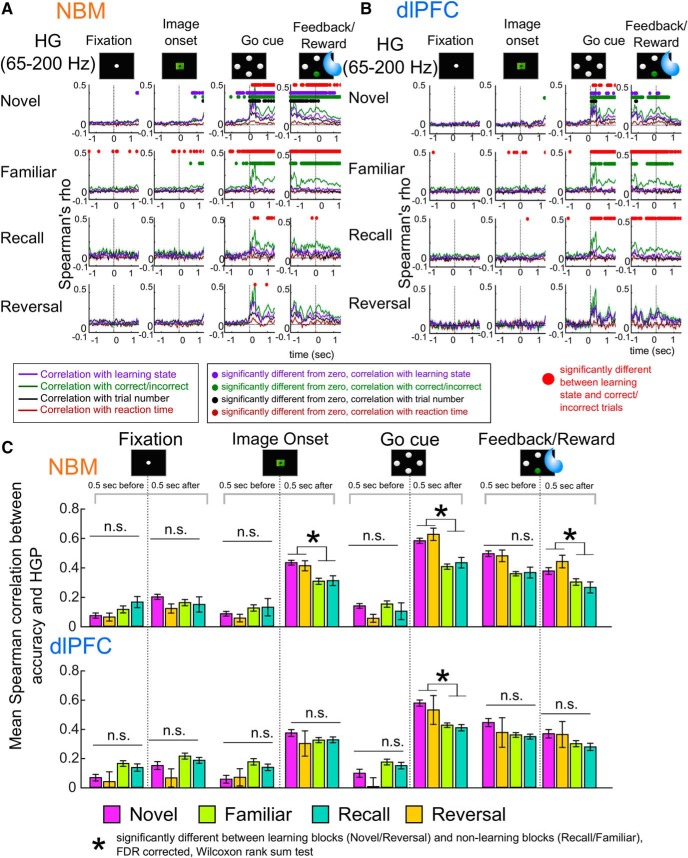
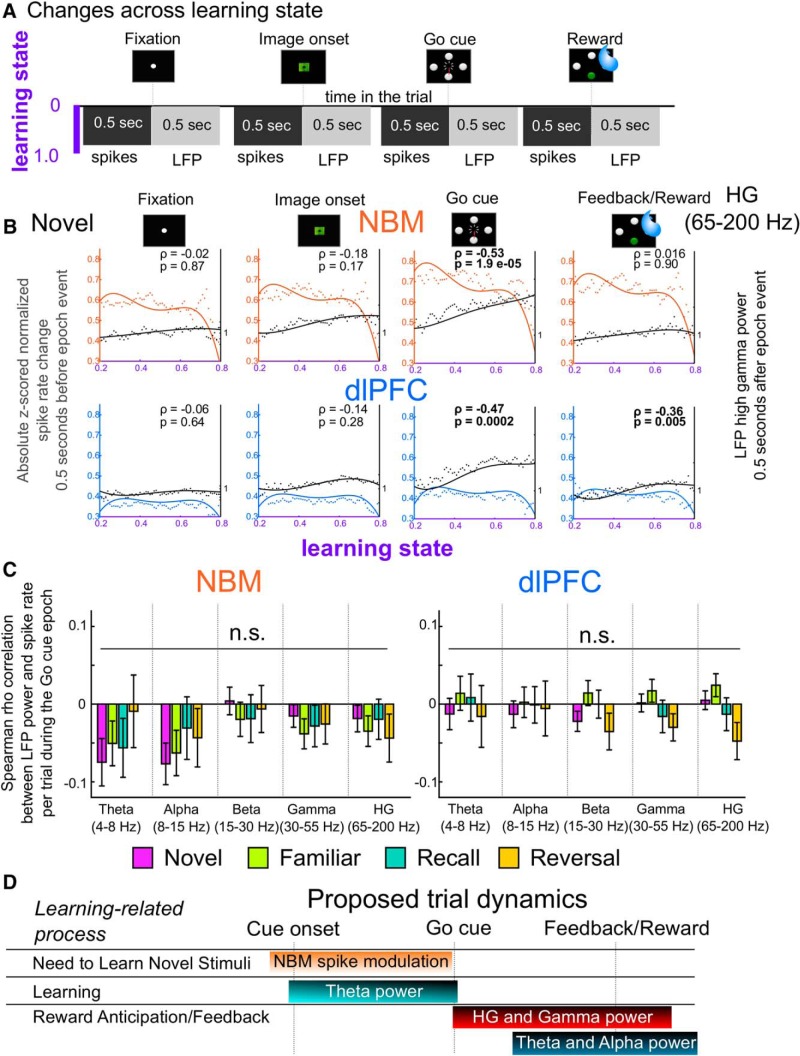
Associative learning is crucial for daily function, involving a complex network of brain regions. One region, the nucleus basalis of Meynert (NBM), is a highly interconnected, largely cholinergic structure implicated in multiple aspects of learning. We show that single neurons in the NBM of nonhuman primates (NHPs; n = 2 males; Macaca mulatta) encode learning a new association through spike rate modulation. However, the power of low-frequency local field potential (LFP) oscillations decreases in response to novel, not-yet-learned stimuli but then increase as learning progresses. Both NBM and the dorsolateral prefrontal cortex encode confidence in novel associations by increasing low- and high-frequency LFP power in anticipation of expected rewards. Finally, NBM high-frequency power dynamics are anticorrelated with spike rate modulations. Therefore, novelty, learning, and reward anticipation are separately encoded through differentiable NBM signals. By signaling both the need to learn and confidence in newly acquired associations, NBM may play a key role in coordinating cortical activity throughout the learning process.SIGNIFICANCE STATEMENT Degradation of cells in a key brain region, the nucleus basalis of Meynert (NBM), correlates with Alzheimer's disease and Parkinson's disease progression. To better understand the role of this brain structure in learning and memory, we examined neural activity in the NBM in behaving nonhuman primates while they performed a learning and memory task. We found that single neurons in NBM encoded both salience and an early learning, or cognitive state, whereas populations of neurons in the NBM and prefrontal cortex encode learned state and reward anticipation. The NBM may thus encode multiple stages of learning. These multimodal signals might be leveraged in future studies to develop neural stimulation to facilitate different stages of learning and memory.
Keywords: encoding cognitive state; learning; local field potential; nucleus basalis of Meynert; single neuron activity.
Copyright © 2018 the authors 0270-6474/18/381942-17$15.00/0.
Figures
Similar articles
-
A New Neural Pathway from the Ventral Striatum to the Nucleus Basalis of Meynert with Functional Implication to Learning and Memory.Mol Neurobiol. 2019 Oct;56(10):7222-7233. doi: 10.1007/s12035-019-1588-0. Epub 2019 Apr 18. Mol Neurobiol. 2019. PMID: 31001802 Free PMC article.
-
Nucleus basalis of Meynert neuronal activity in Parkinson's disease.J Neurosurg. 2019 Feb 22;132(2):574-582. doi: 10.3171/2018.11.JNS182386. Print 2020 Feb 1. J Neurosurg. 2019. PMID: 30797189
-
Nucleus basalis of Meynert modulates signal processing in rat layer 5 somatosensory cortex but leads to memory impairment and tactile discrimination deficits following lesion.Behav Brain Res. 2020 May 27;386:112608. doi: 10.1016/j.bbr.2020.112608. Epub 2020 Mar 16. Behav Brain Res. 2020. PMID: 32194192
-
The Nucleus Basalis of Meynert and Its Role in Deep Brain Stimulation for Cognitive Disorders: A Historical Perspective.J Alzheimers Dis. 2019;69(4):905-919. doi: 10.3233/JAD-180133. J Alzheimers Dis. 2019. PMID: 31104014 Review.
-
Deep Brain Stimulation in Alzheimer's Disease: Targeting the Nucleus Basalis of Meynert.J Alzheimers Dis. 2021;80(1):53-70. doi: 10.3233/JAD-201141. J Alzheimers Dis. 2021. PMID: 33492288 Review.
Cited by
-
The neural circuits of monogamous behavior.Front Neural Circuits. 2022 Sep 29;16:978344. doi: 10.3389/fncir.2022.978344. eCollection 2022. Front Neural Circuits. 2022. PMID: 36247729 Free PMC article. Review.
-
Forebrain Cholinergic Signaling: Wired and Phasic, Not Tonic, and Causing Behavior.J Neurosci. 2020 Jan 22;40(4):712-719. doi: 10.1523/JNEUROSCI.1305-19.2019. J Neurosci. 2020. PMID: 31969489 Free PMC article. Review.
-
Metric magnetic resonance imaging analysis reveals pronounced substantia-innominata atrophy in dementia with Lewy bodies with a psychiatric onset.Front Aging Neurosci. 2022 Oct 5;14:815813. doi: 10.3389/fnagi.2022.815813. eCollection 2022. Front Aging Neurosci. 2022. PMID: 36274999 Free PMC article.
-
The role of the nucleus basalis of Meynert in neuromodulation therapy: a systematic review from the perspective of neural network oscillations.Front Aging Neurosci. 2024 Apr 8;16:1376764. doi: 10.3389/fnagi.2024.1376764. eCollection 2024. Front Aging Neurosci. 2024. PMID: 38650866 Free PMC article. Review.
-
Role of Cholinergic Signaling in Alzheimer's Disease.Molecules. 2022 Mar 10;27(6):1816. doi: 10.3390/molecules27061816. Molecules. 2022. PMID: 35335180 Free PMC article. Review.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources