

Toward dynamic structural biology: Two decades of single-molecule Förster resonance energy transfer
- PMID: 29348210
- PMCID: PMC6200918
- DOI: 10.1126/science.aan1133
Toward dynamic structural biology: Two decades of single-molecule Förster resonance energy transfer
Abstract
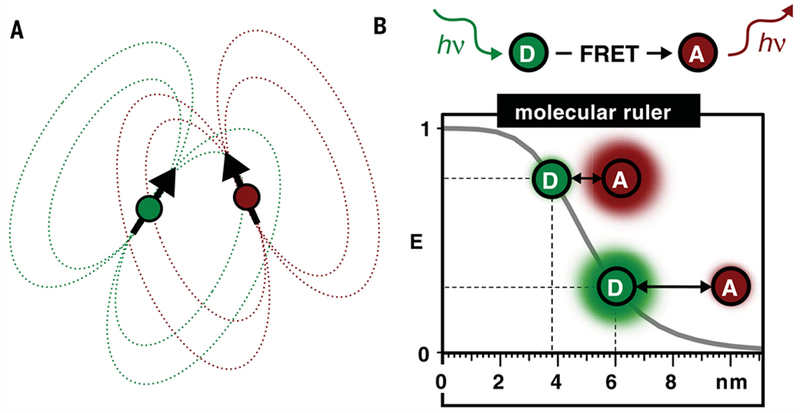
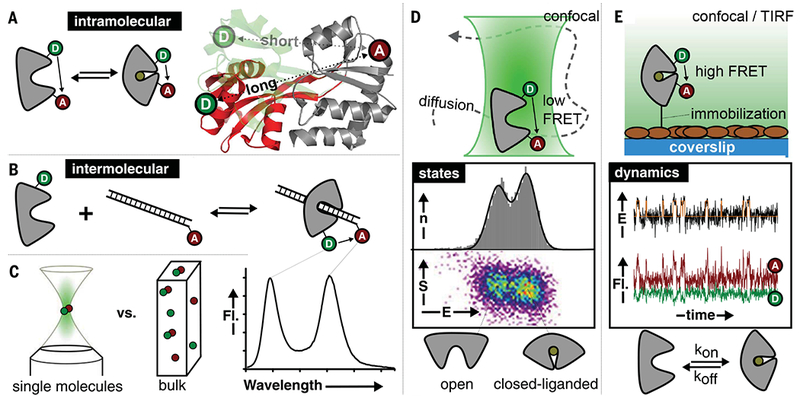
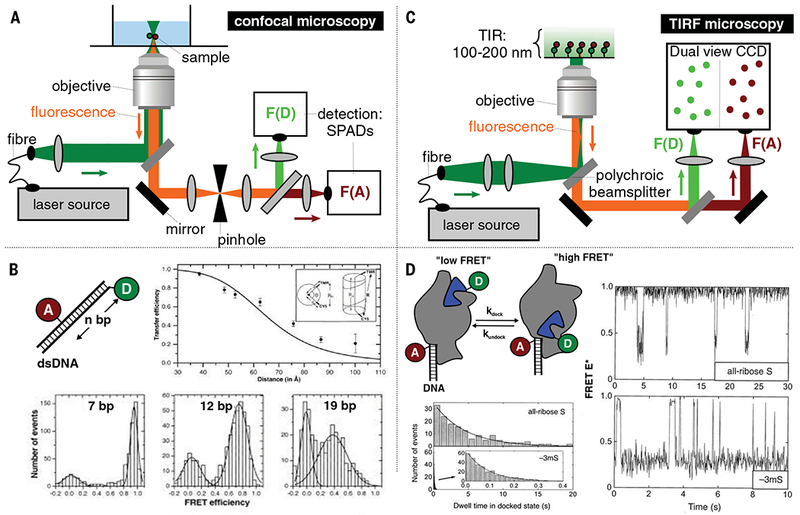
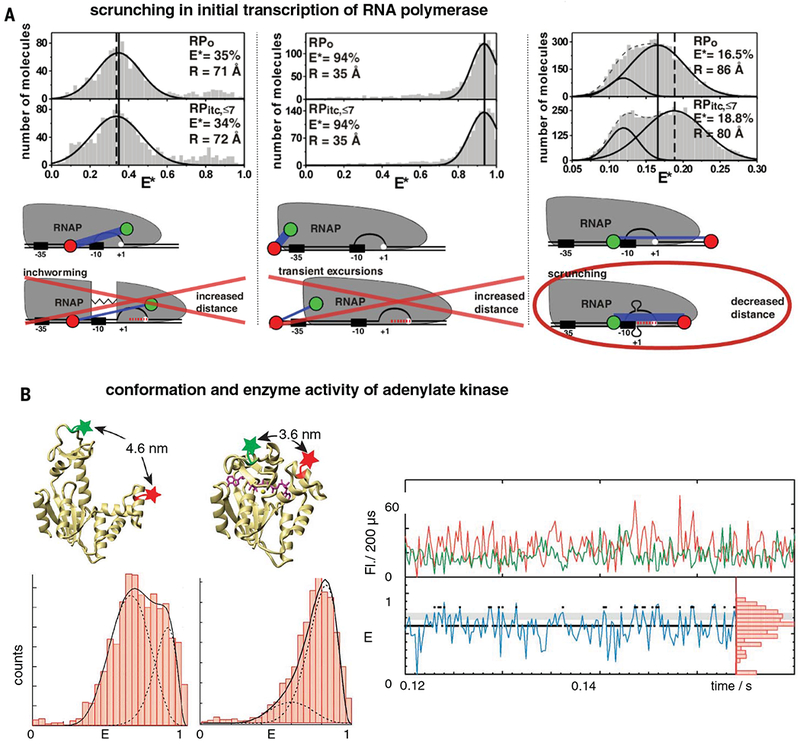
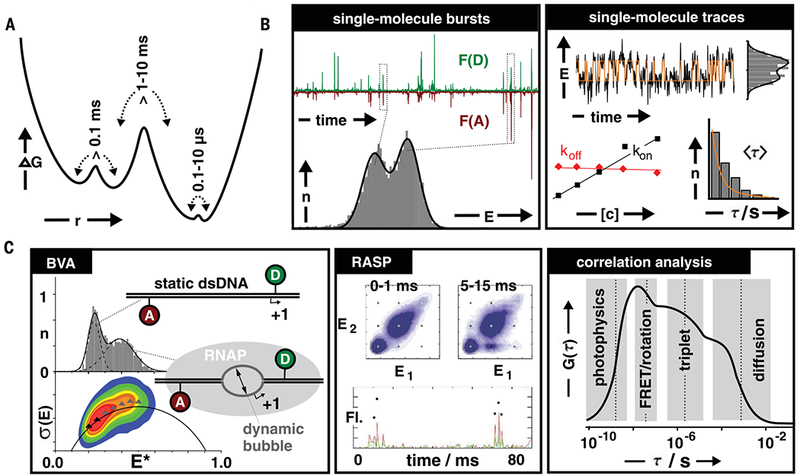
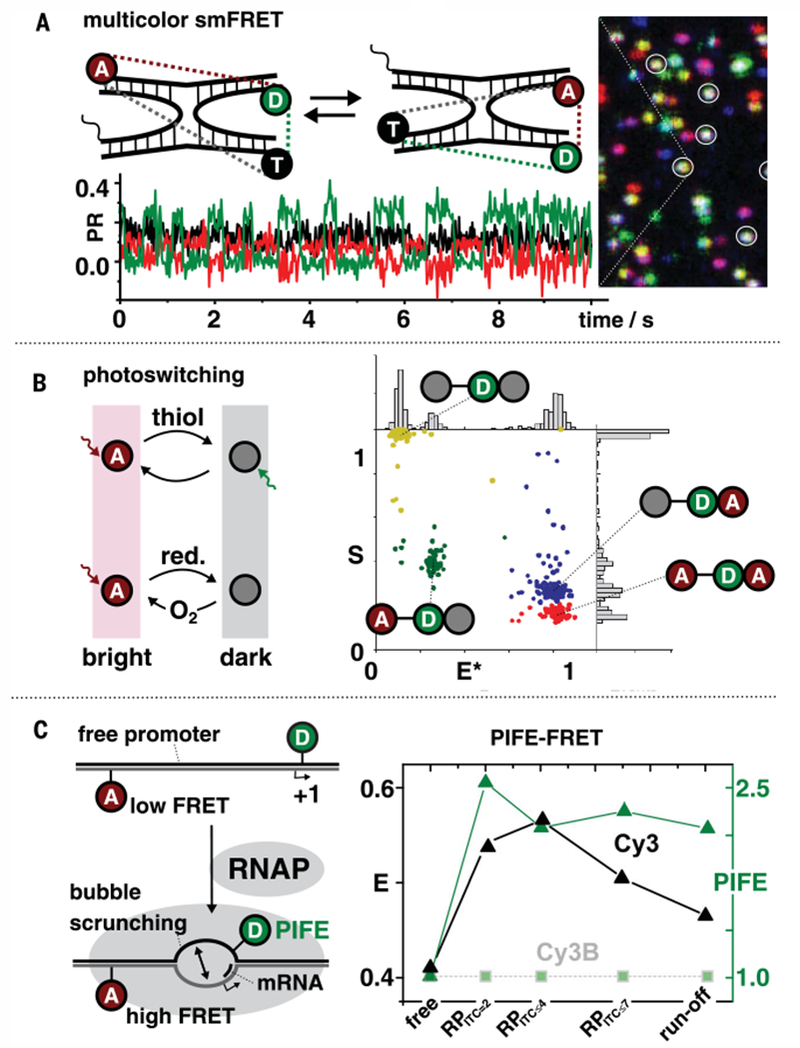
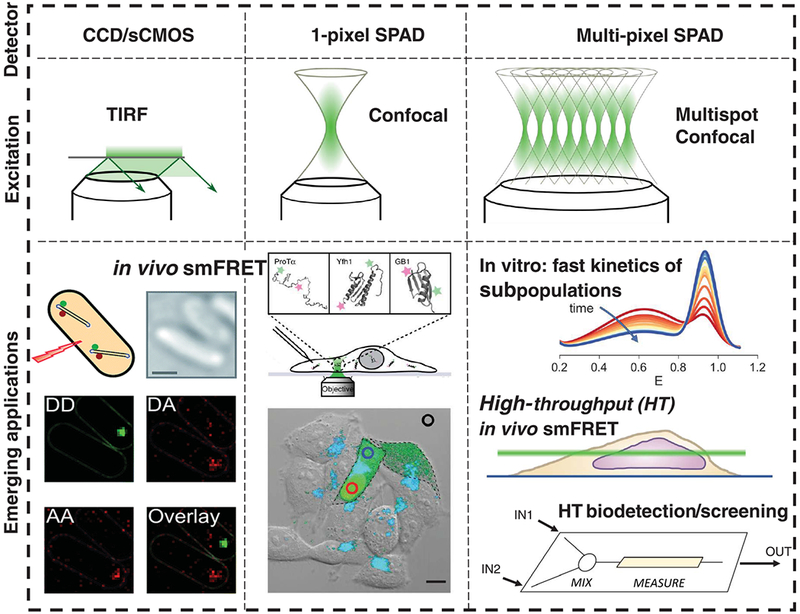
Classical structural biology can only provide static snapshots of biomacromolecules. Single-molecule Förster resonance energy transfer (smFRET) paved the way for studying dynamics in macromolecular structures under biologically relevant conditions. Since its first implementation in 1996, smFRET experiments have confirmed previously hypothesized mechanisms and provided new insights into many fundamental biological processes, such as DNA maintenance and repair, transcription, translation, and membrane transport. We review 22 years of contributions of smFRET to our understanding of basic mechanisms in biochemistry, molecular biology, and structural biology. Additionally, building on current state-of-the-art implementations of smFRET, we highlight possible future directions for smFRET in applications such as biosensing, high-throughput screening, and molecular diagnostics.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
References
-
- Brünger AT, X-ray crystallography and NMR reveal complementary views of structure and dynamics. Nat. Struct. Biol 4 (suppl.), 862–865 (1997). pmid: - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
