

A pathway for mitotic chromosome formation
- PMID: 29348367
- PMCID: PMC5924687
- DOI: 10.1126/science.aao6135
A pathway for mitotic chromosome formation
Abstract
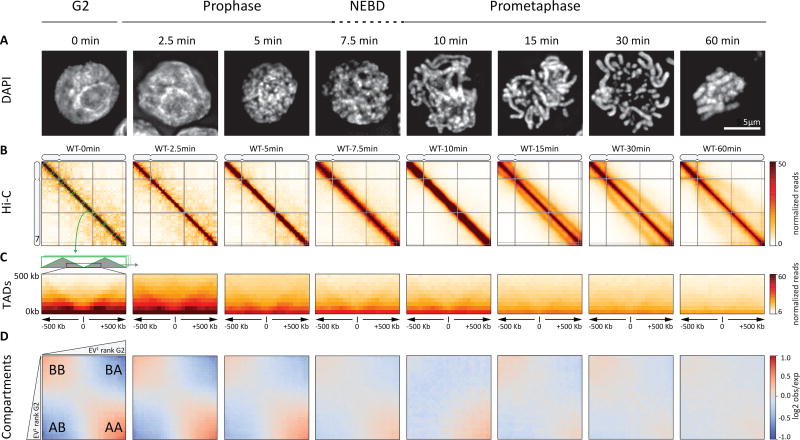
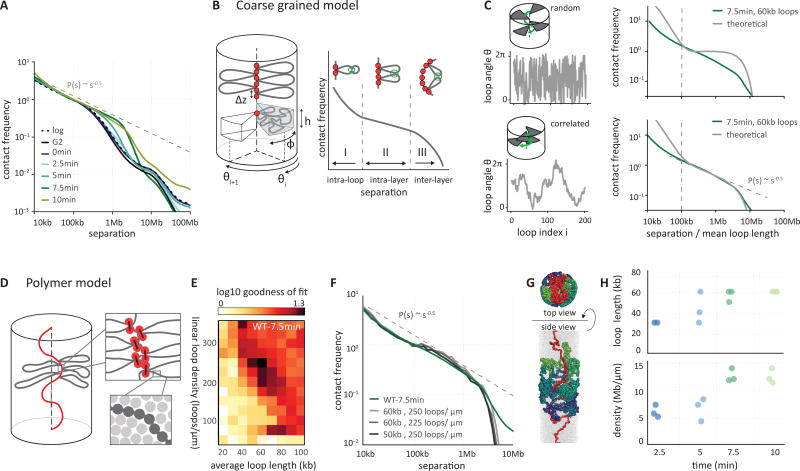
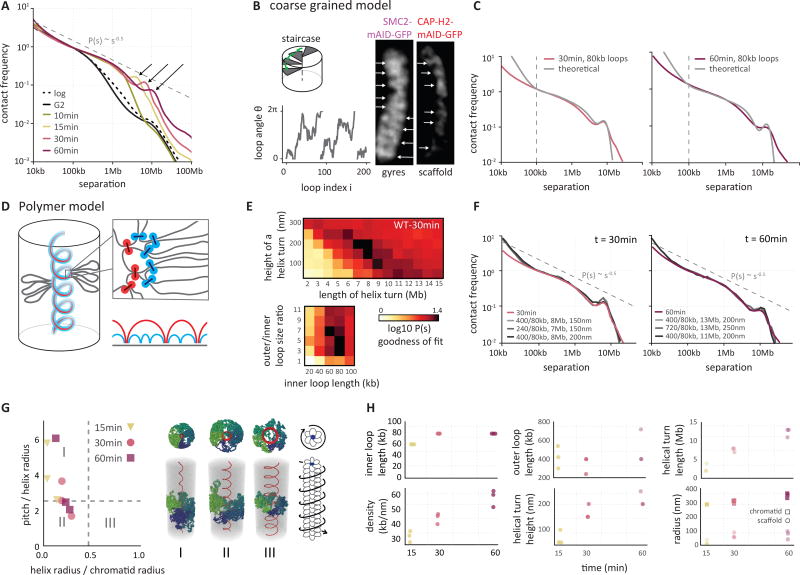
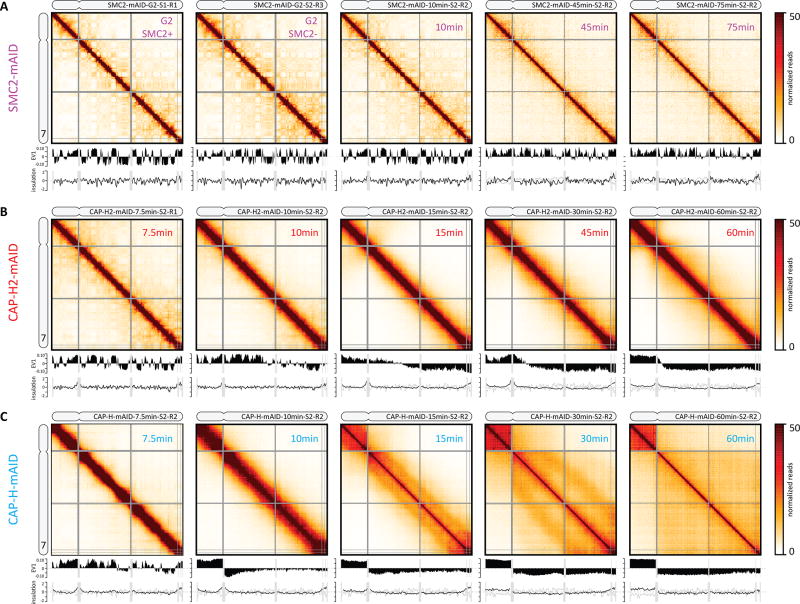
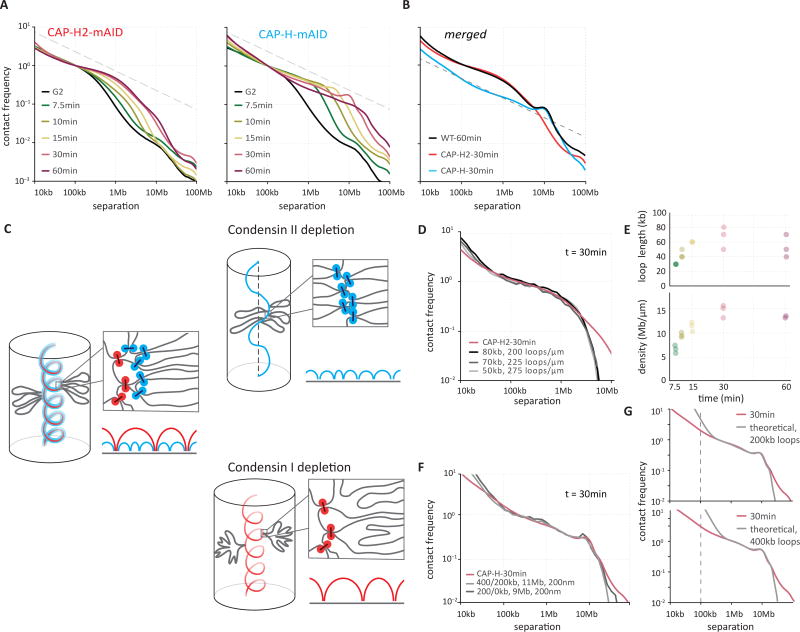
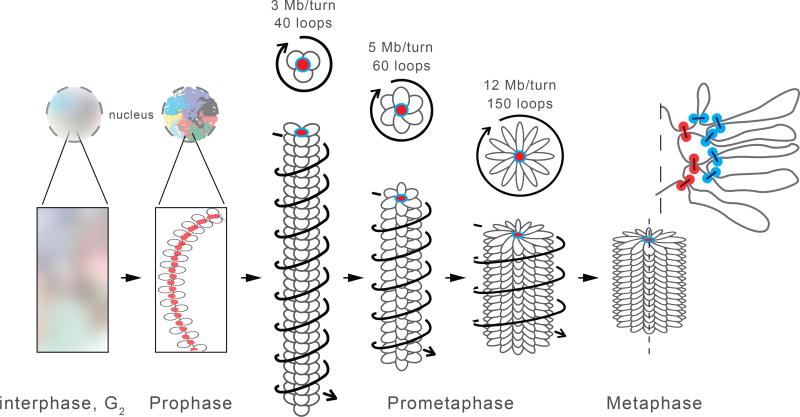
Mitotic chromosomes fold as compact arrays of chromatin loops. To identify the pathway of mitotic chromosome formation, we combined imaging and Hi-C analysis of synchronous DT40 cell cultures with polymer simulations. Here we show that in prophase, the interphase organization is rapidly lost in a condensin-dependent manner, and arrays of consecutive 60-kilobase (kb) loops are formed. During prometaphase, ~80-kb inner loops are nested within ~400-kb outer loops. The loop array acquires a helical arrangement with consecutive loops emanating from a central "spiral staircase" condensin scaffold. The size of helical turns progressively increases to ~12 megabases during prometaphase. Acute depletion of condensin I or II shows that nested loops form by differential action of the two condensins, whereas condensin II is required for helical winding.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
Comment in
-
Chromosome biology: A stairway to mitotic chromosome assembly.Nat Rev Mol Cell Biol. 2018 Feb 20;19(3):139. doi: 10.1038/nrm.2018.13. Nat Rev Mol Cell Biol. 2018. PMID: 29459658 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
