

Myelination of Purkinje axons is critical for resilient synaptic transmission in the deep cerebellar nucleus
- PMID: 29348594
- PMCID: PMC5773691
- DOI: 10.1038/s41598-018-19314-0
Myelination of Purkinje axons is critical for resilient synaptic transmission in the deep cerebellar nucleus
Abstract
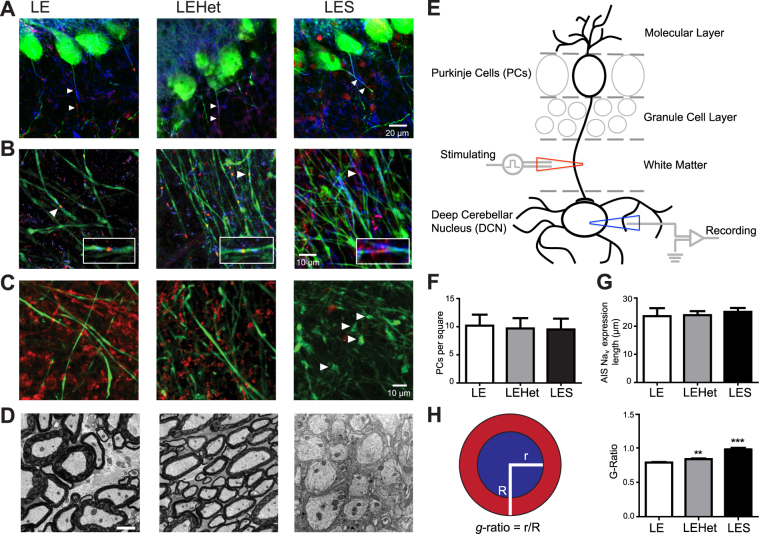
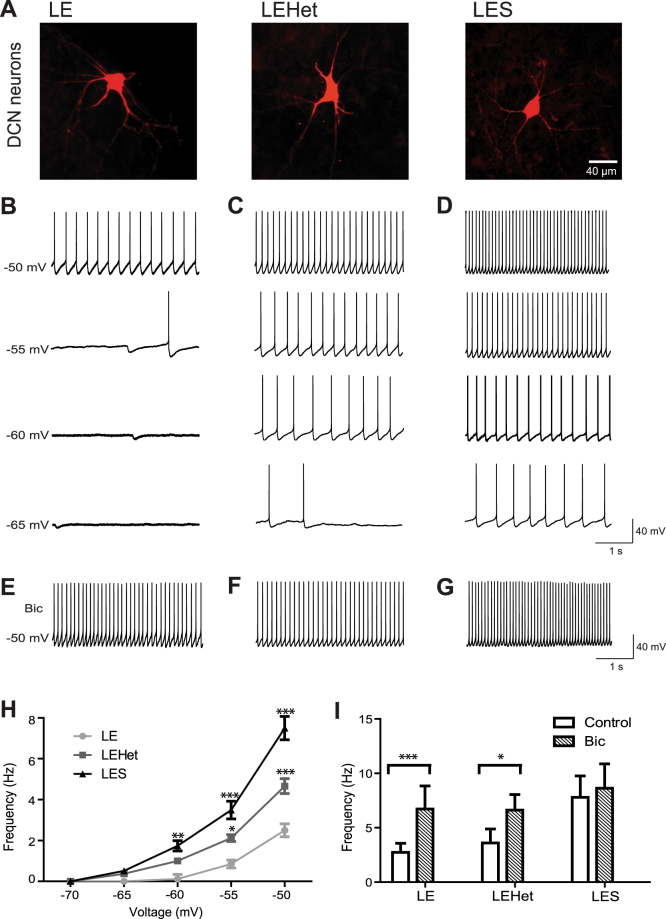
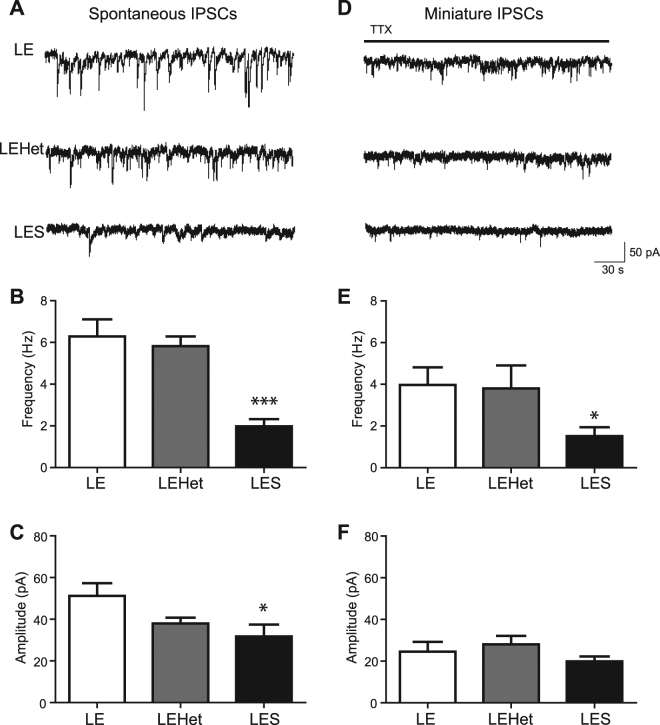
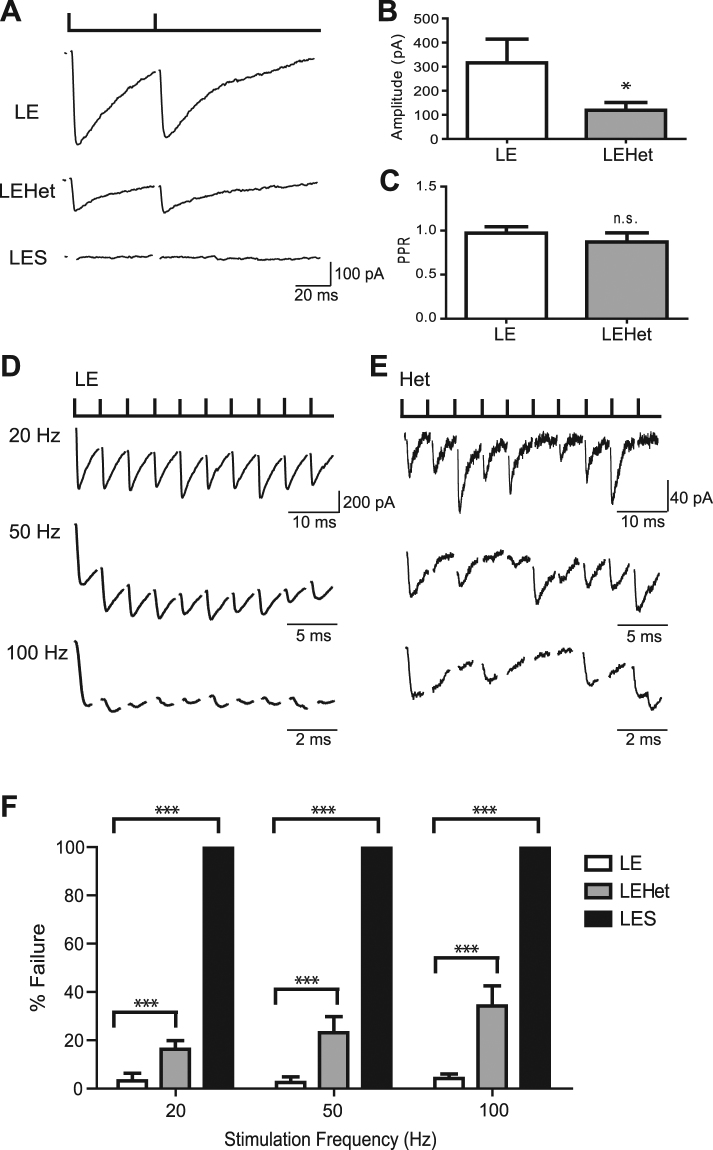
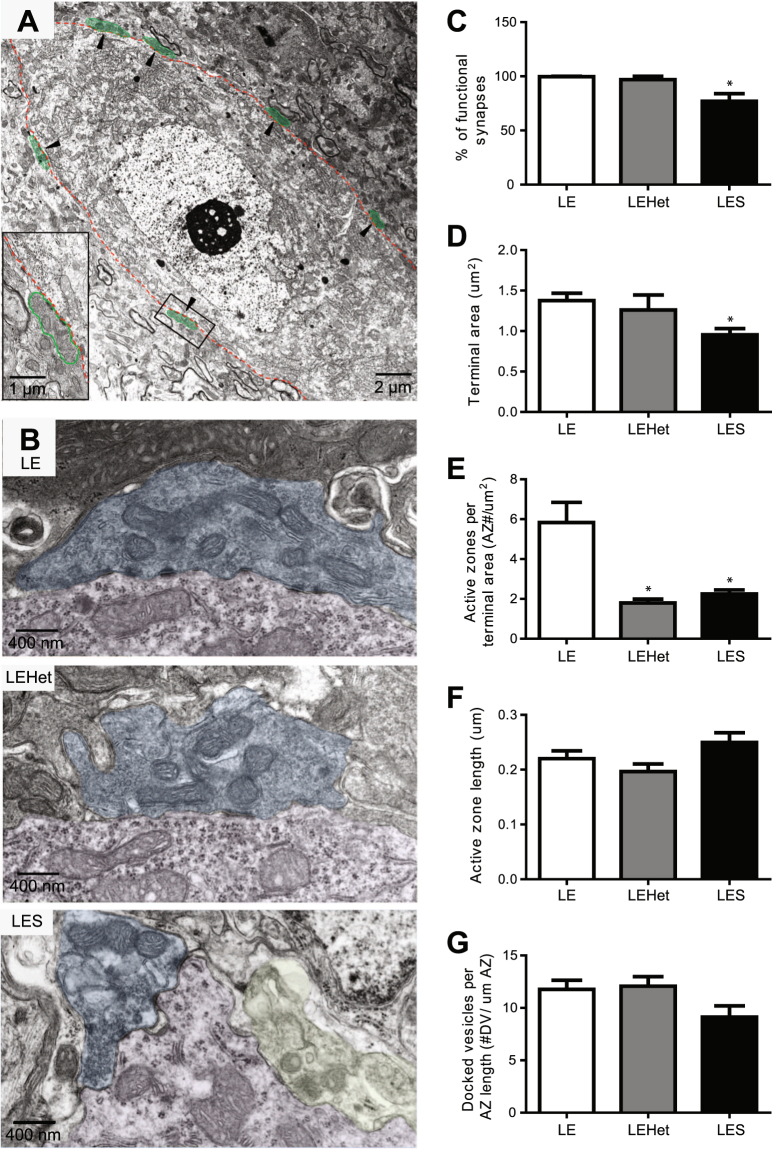
The roles of myelin in maintaining axonal integrity and action potential (AP) propagation are well established, but its role in synapse maintenance and neurotransmission remains largely understudied. Here, we investigated how Purkinje axon myelination regulates synaptic transmission in the Purkinje to deep cerebellar nuclei (DCN) synapses using the Long Evans Shaker (LES) rat, which lacks compact myelin and thus displays severe locomotion deficits. DCN neurons fired spontaneous action potentials (APs), whose frequencies were dependent on the extent of myelin. In the LES cerebellum with severe myelin deficiency, DCN neurons were hyper-excitable, exhibiting spontaneous AP firing at a much higher frequency compared to those from wild type (LE) and heterozygote (LEHet) rats. The hyper-excitability in LES DCN neurons resulted from reduced inhibitory GABAergic inputs from Purkinje cells to DCN neurons. Corresponding with functional alterations including failures of AP propagation, electron microscopic analysis revealed anatomically fewer active zones at the presynaptic terminals of Purkinje cells in both LEHet and LES rats. Taken together, these studies suggest that proper axonal myelination critically regulates presynaptic terminal structure and function and directly impacts synaptic transmission in the Purkinje cell-DCN cell synapse in the cerebellum.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
