

Syringic acid, a phenolic acid, promotes osteoblast differentiation by stimulation of Runx2 expression and targeting of Smad7 by miR-21 in mouse mesenchymal stem cells
- PMID: 29350343
- PMCID: PMC6039342
- DOI: 10.1007/s12079-018-0449-3
Syringic acid, a phenolic acid, promotes osteoblast differentiation by stimulation of Runx2 expression and targeting of Smad7 by miR-21 in mouse mesenchymal stem cells
Abstract
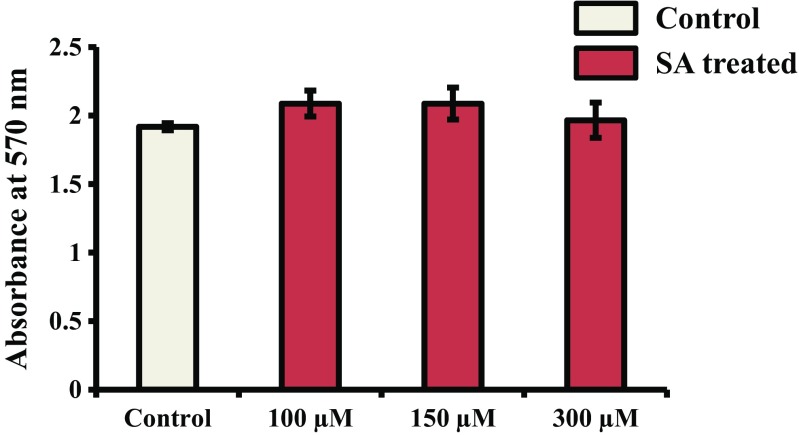
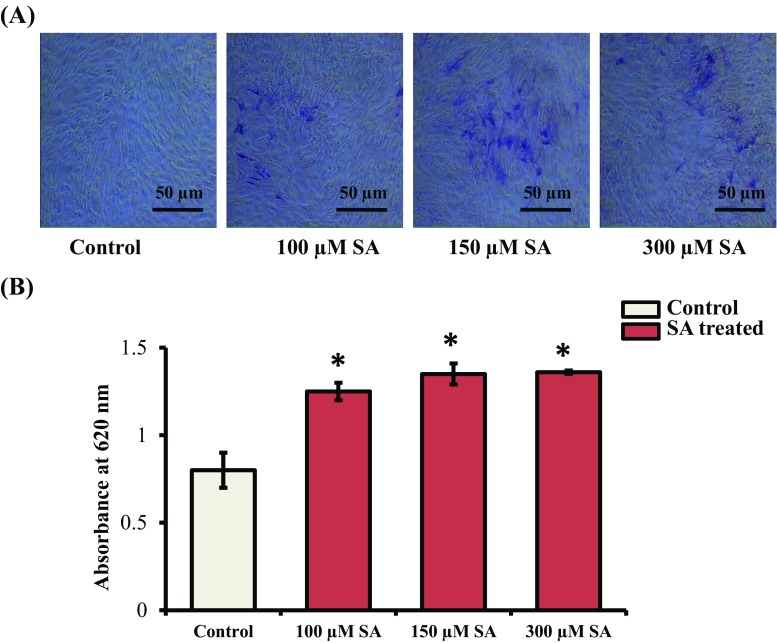
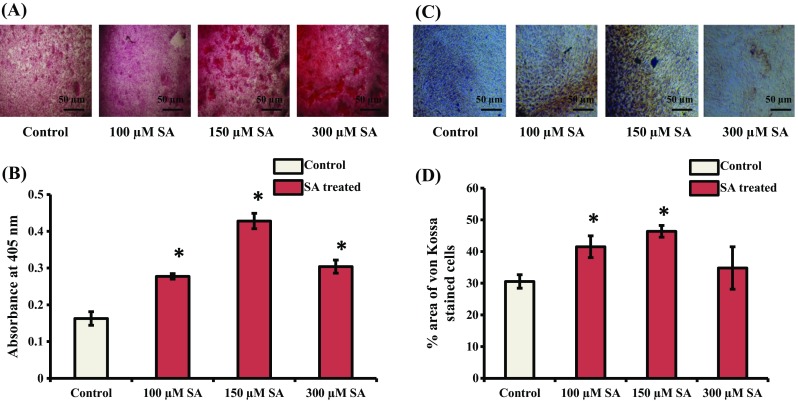
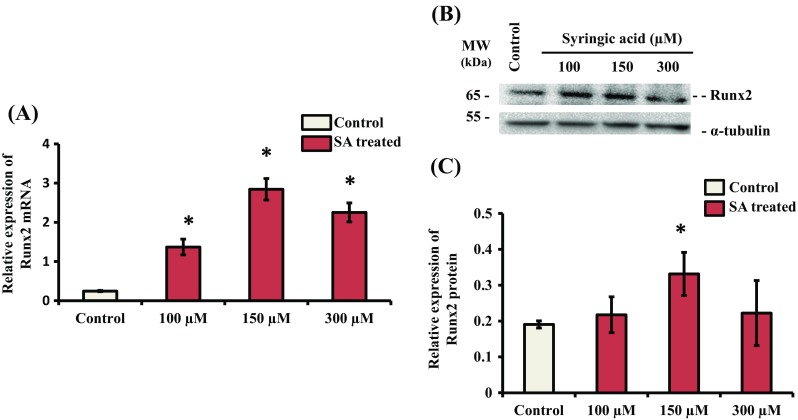
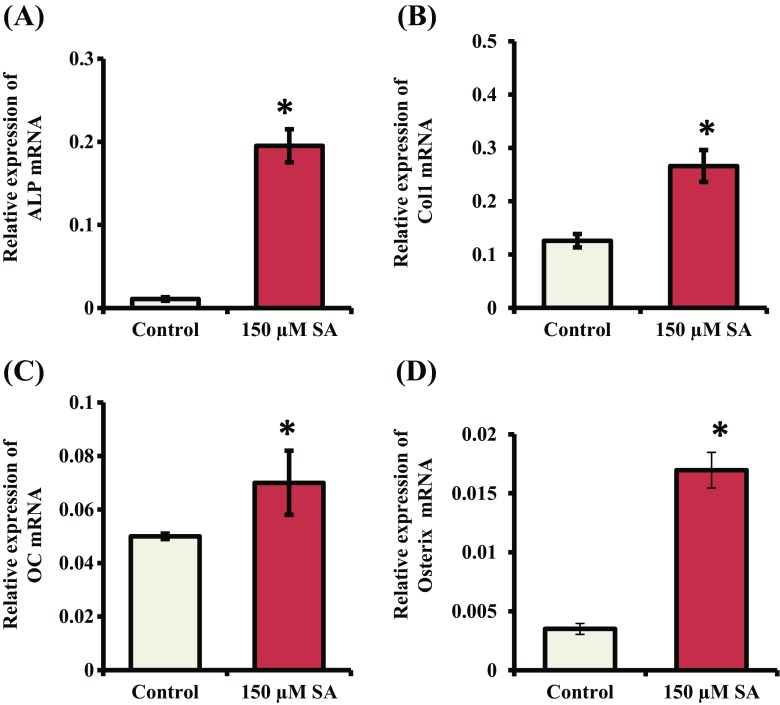
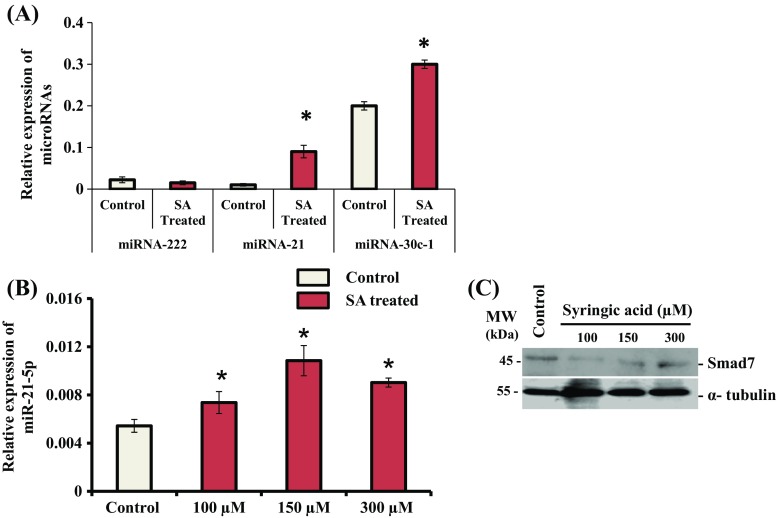
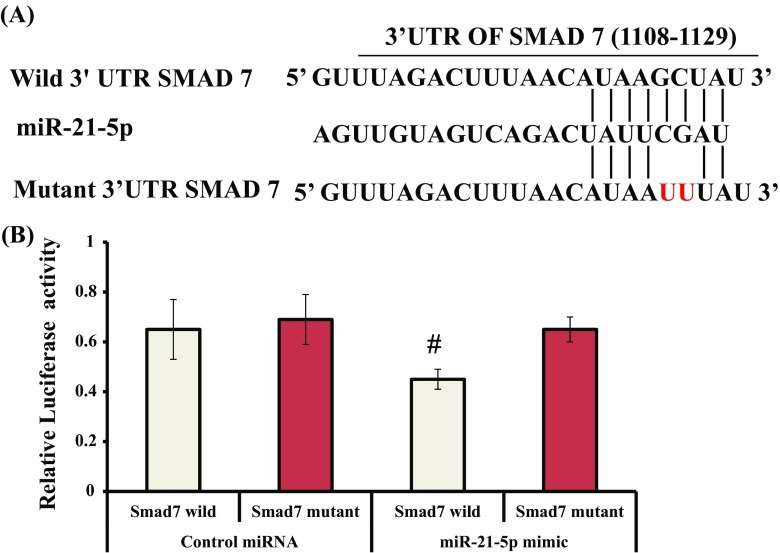
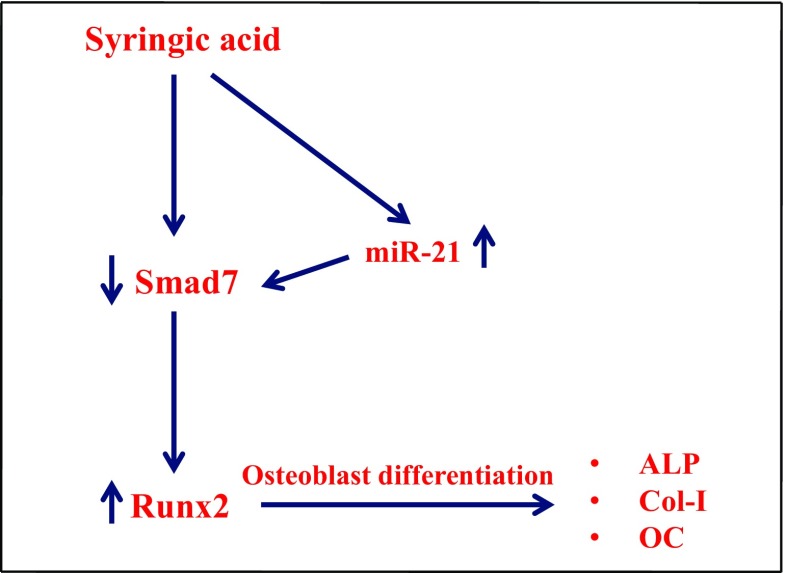
Syringic acid (SA), a phenolic acid, has been used in Chinese and Indian medicine for treating diabetes but its role in osteogenesis has not yet been investigated. In the present study, at the molecular and cellular levels, we evaluated the effects of SA on osteoblast differentiation. At the cellular level, there was increased alkaline phosphatase (ALP) activity and calcium deposition by SA treatment in mouse mesenchymal stem cells (mMSCs). At the molecular level, SA treatment of these cells stimulated expression of Runx2, a bone transcription factor, and of osteoblast differentiation marker genes such as ALP, type I collagen, and osteocalcin. It is known that Smad7 is an antagonist of TGF-β/Smad signaling and is a negative regulator of Runx2. microRNAs (miRNAs) play a key role in the regulation of osteogenesis genes at the post-transcriptional level and studies have reported that Smad7 is one of the target genes of miR-21. We found that there was down regulation of Smad7 and up regulation of miR-21 in SA-treated mMSCs. We further identified that the 3'-untranslated region (UTR) of Smad7 was directly targeted by miR-21 in these cells. Thus, our results suggested that SA promotes osteoblast differentiation via increased expression of Runx2 by miR-21-mediated down regulation of Smad7. Hence, SA may have potential in orthopedic applications.
Keywords: Bone; Runx2; Smad7; Syringic acid; miR-21.
Figures


References
-
- Arumugam B, Vairamani M, Partridge NC, Selvamurugan N (2017) Characterization of Runx2 phosphorylation sites required for TGF-β1-mediated stimulation of matrix Metalloproteinase-13 expression in osteoblastic cells. J Cell Physiol 10.1002/jcp.25964 - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
