

A Membrane G-Protein-Coupled Estrogen Receptor Is Necessary but Not Sufficient for Sex Differences in Zebra Finch Auditory Coding
- PMID: 29351614
- PMCID: PMC5839738
- DOI: 10.1210/en.2017-03102
A Membrane G-Protein-Coupled Estrogen Receptor Is Necessary but Not Sufficient for Sex Differences in Zebra Finch Auditory Coding
Abstract
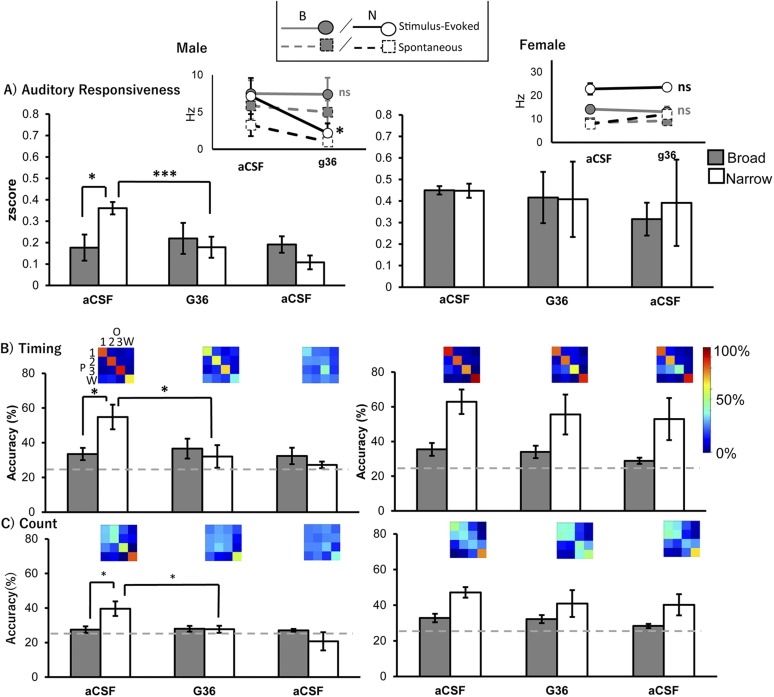
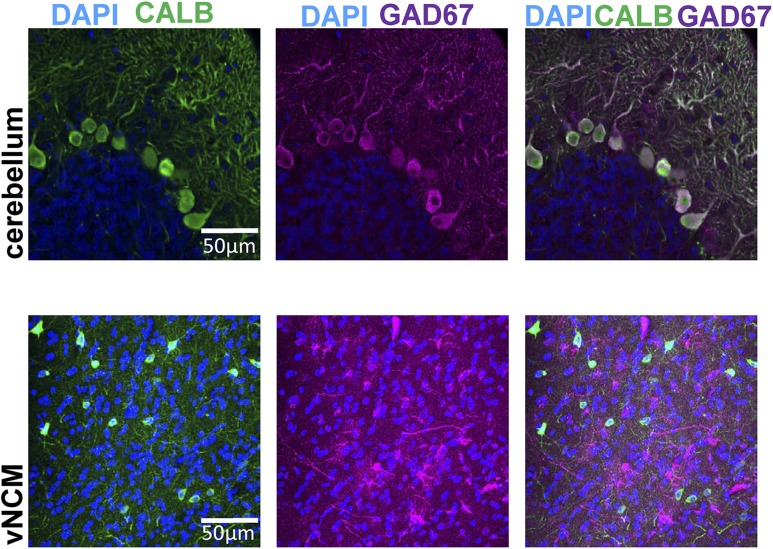
Estradiol acts as a neuromodulator in brain regions important for cognition and sensory processing. Estradiol also shapes brain sex differences but rarely have these concepts been considered simultaneously. In male and female songbirds, estradiol rapidly increases within the auditory forebrain during song exposure and enhances local auditory processing. We tested whether G-protein-coupled estrogen receptor 1 (GPER1), a membrane-bound estrogen receptor, is necessary and sufficient for neuroestrogen regulation of forebrain auditory processing in male and female zebra finches (Taeniopygia guttata). At baseline, we observed that females had elevated single-neuron responses to songs vs males. In males, narrow-spiking (NS) neurons were more responsive to conspecific songs than broad-spiking (BS) neurons, yet cell types were similarly auditory responsive in females. Following acute inactivation of GPER1, auditory responsiveness and coding were suppressed in male NS yet unchanged in female NS and in BS of both sexes. By contrast, GPER1 activation did not mimic previously established estradiol actions in either sex. Lastly, the expression of GPER1 and its coexpression with an inhibitory neuron marker were similarly abundant in males and females, confirming anatomical similarity in the auditory forebrain. In this study, we found: (1) a role for GPER1 in regulating sensory processing and (2) a sex difference in auditory processing of complex vocalizations in a cell type-specific manner. These results reveal sex specificity of a rapid estrogen signaling mechanism in which neuromodulation accounts and/or compensates for brain sex differences, dependent on cell type, in brain regions that are anatomically similar in both sexes.
Conflict of interest statement
The authors have nothing to disclose.
Figures






Similar articles
-
Acute neuroestrogen blockade attenuates song-induced immediate early gene expression in auditory regions of male and female zebra finches.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2020 Jan;206(1):15-31. doi: 10.1007/s00359-019-01382-w. Epub 2019 Nov 28. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2020. PMID: 31781892 Free PMC article.
-
Norepinephrine enhances song responsiveness and encoding in the auditory forebrain of male zebra finches.J Neurophysiol. 2018 Jan 1;119(1):209-220. doi: 10.1152/jn.00251.2017. Epub 2017 Oct 11. J Neurophysiol. 2018. PMID: 29021389 Free PMC article.
-
Developmental experience alters information coding in auditory midbrain and forebrain neurons.Dev Neurobiol. 2010 Mar;70(4):235-52. doi: 10.1002/dneu.20783. Dev Neurobiol. 2010. PMID: 20039264 Free PMC article.
-
Sex differences and rapid estrogen signaling: A look at songbird audition.Front Neuroendocrinol. 2015 Jul;38:37-49. doi: 10.1016/j.yfrne.2015.01.001. Epub 2015 Jan 28. Front Neuroendocrinol. 2015. PMID: 25637753 Free PMC article. Review.
-
It takes a seasoned bird to be a good listener: communication between the sexes.Curr Opin Neurobiol. 2016 Jun;38:12-7. doi: 10.1016/j.conb.2016.01.005. Epub 2016 Jan 25. Curr Opin Neurobiol. 2016. PMID: 26820470 Free PMC article. Review.
Cited by
-
Life-Stage Dependent Plasticity in the Auditory System of a Songbird Is Signal and Emitter-Specific.Front Neurosci. 2020 Dec 4;14:588672. doi: 10.3389/fnins.2020.588672. eCollection 2020. Front Neurosci. 2020. PMID: 33343284 Free PMC article.
-
Estrogens rapidly shape synaptic and intrinsic properties to regulate the temporal precision of songbird auditory neurons.Cereb Cortex. 2023 Mar 21;33(7):3401-3420. doi: 10.1093/cercor/bhac280. Cereb Cortex. 2023. PMID: 35849820 Free PMC article.
-
Temporal and bidirectional influences of estradiol on voluntary wheel running in adult female and male rats.Horm Behav. 2020 Apr;120:104694. doi: 10.1016/j.yhbeh.2020.104694. Epub 2020 Jan 27. Horm Behav. 2020. PMID: 31978389 Free PMC article.
-
Adult-like neural representation of species-specific songs in the auditory forebrain of zebra finch nestlings.Dev Neurobiol. 2021 Mar;81(2):123-138. doi: 10.1002/dneu.22802. Epub 2021 Jan 29. Dev Neurobiol. 2021. PMID: 33369121 Free PMC article.
-
Auditory learning in an operant task with social reinforcement is dependent on neuroestrogen synthesis in the male songbird auditory cortex.Horm Behav. 2020 May;121:104713. doi: 10.1016/j.yhbeh.2020.104713. Epub 2020 Feb 19. Horm Behav. 2020. PMID: 32057821 Free PMC article.
References
-
- Nottebohm F, Arnold AP. Sexual dimorphism in vocal control areas of the songbird brain. Science. 1976;194(4261):211–213. - PubMed
-
- Hamaide J, De Groof G, Van Steenkiste G, Jeurissen B, Van Audekerke J, Naeyaert M, Van Ruijssevelt L, Cornil C, Sijbers J, Verhoye M, Van der Linden A. Exploring sex differences in the adult zebra finch brain: in vivo diffusion tensor imaging and ex vivo super-resolution track density imaging. Neuroimage. 2017;146:789–803. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
