

Spatiotemporal organization of exocytosis emerges during neuronal shape change
- PMID: 29351997
- PMCID: PMC5839795
- DOI: 10.1083/jcb.201709064
Spatiotemporal organization of exocytosis emerges during neuronal shape change
Abstract
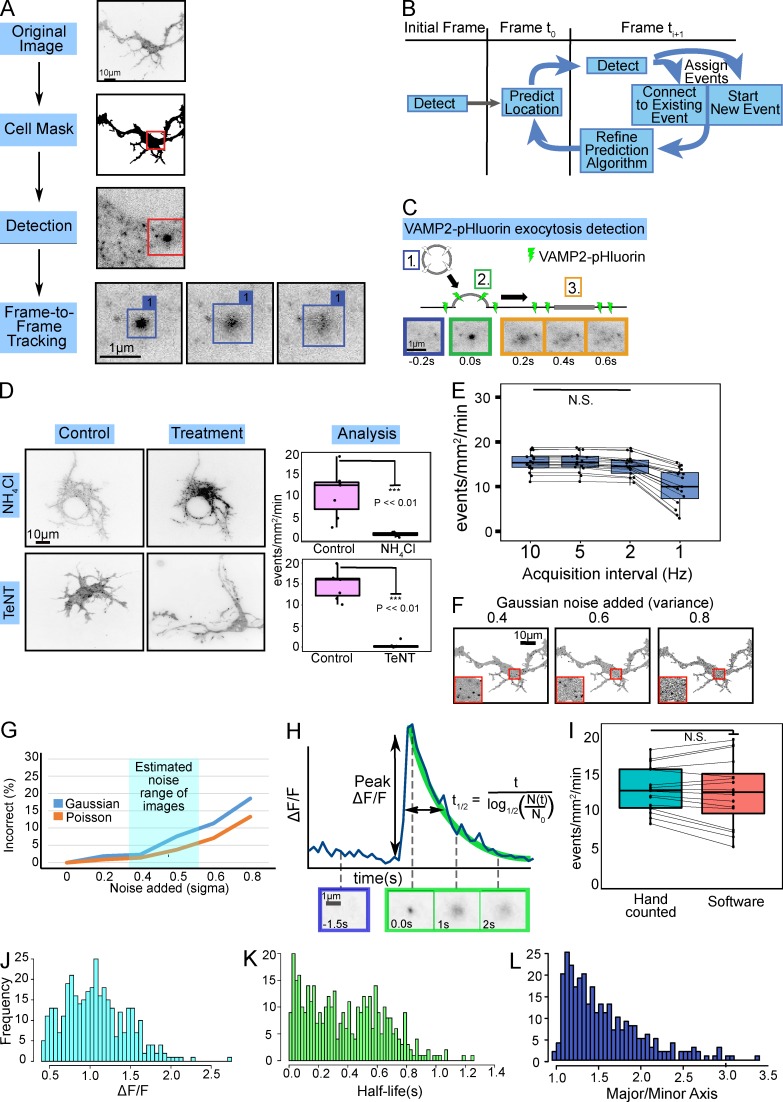
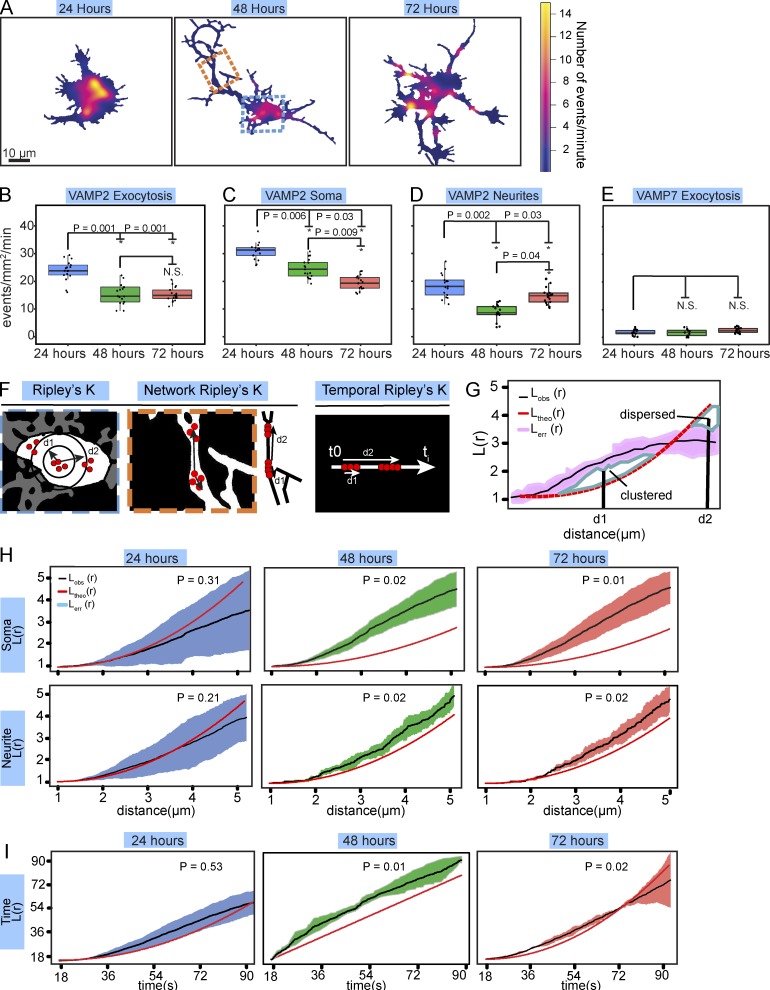
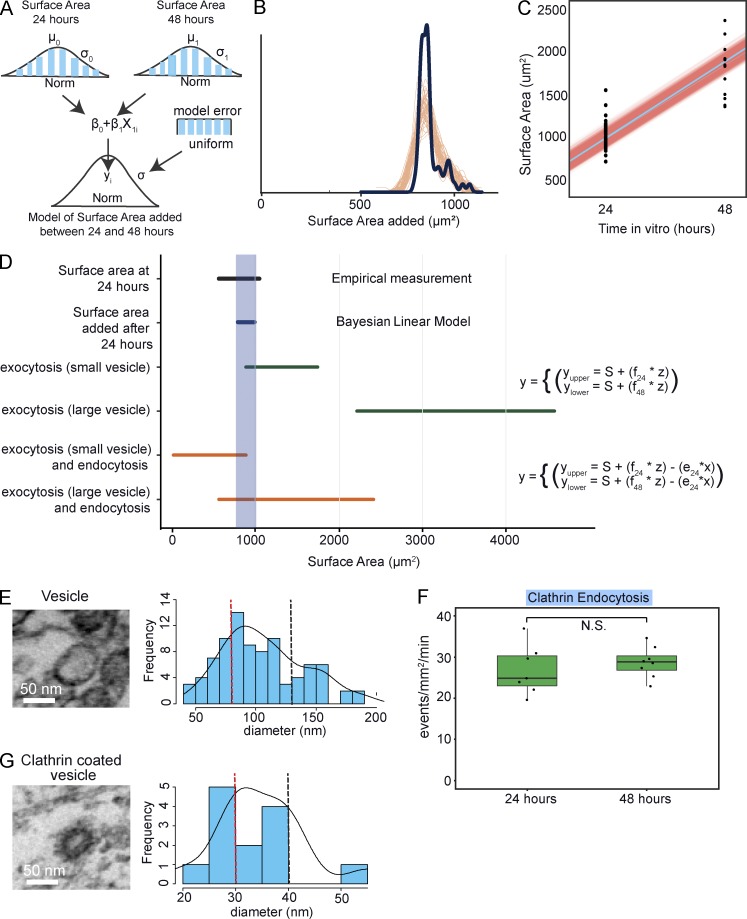
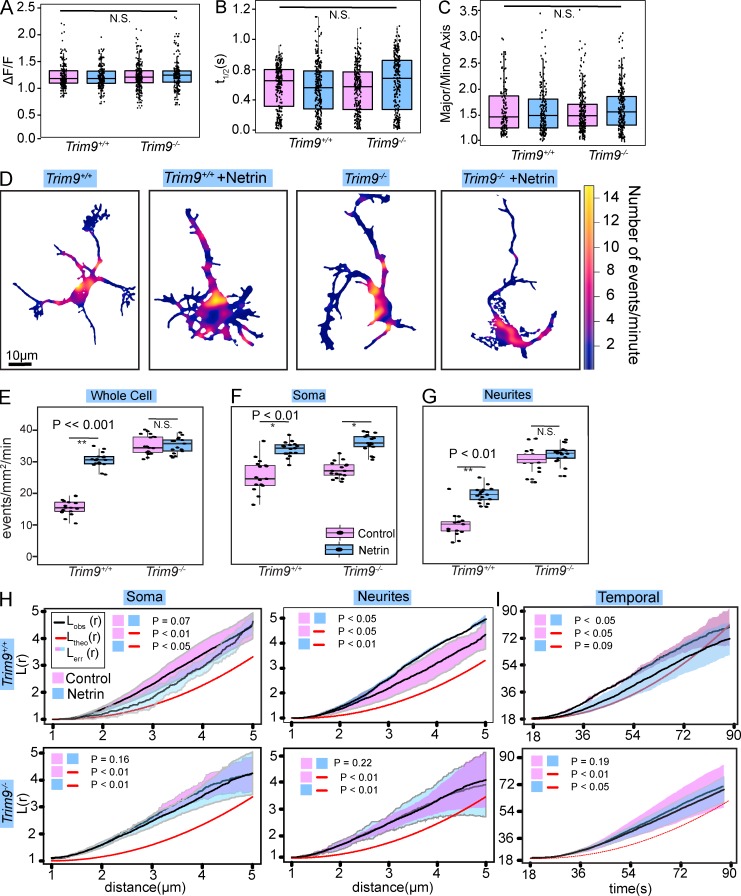
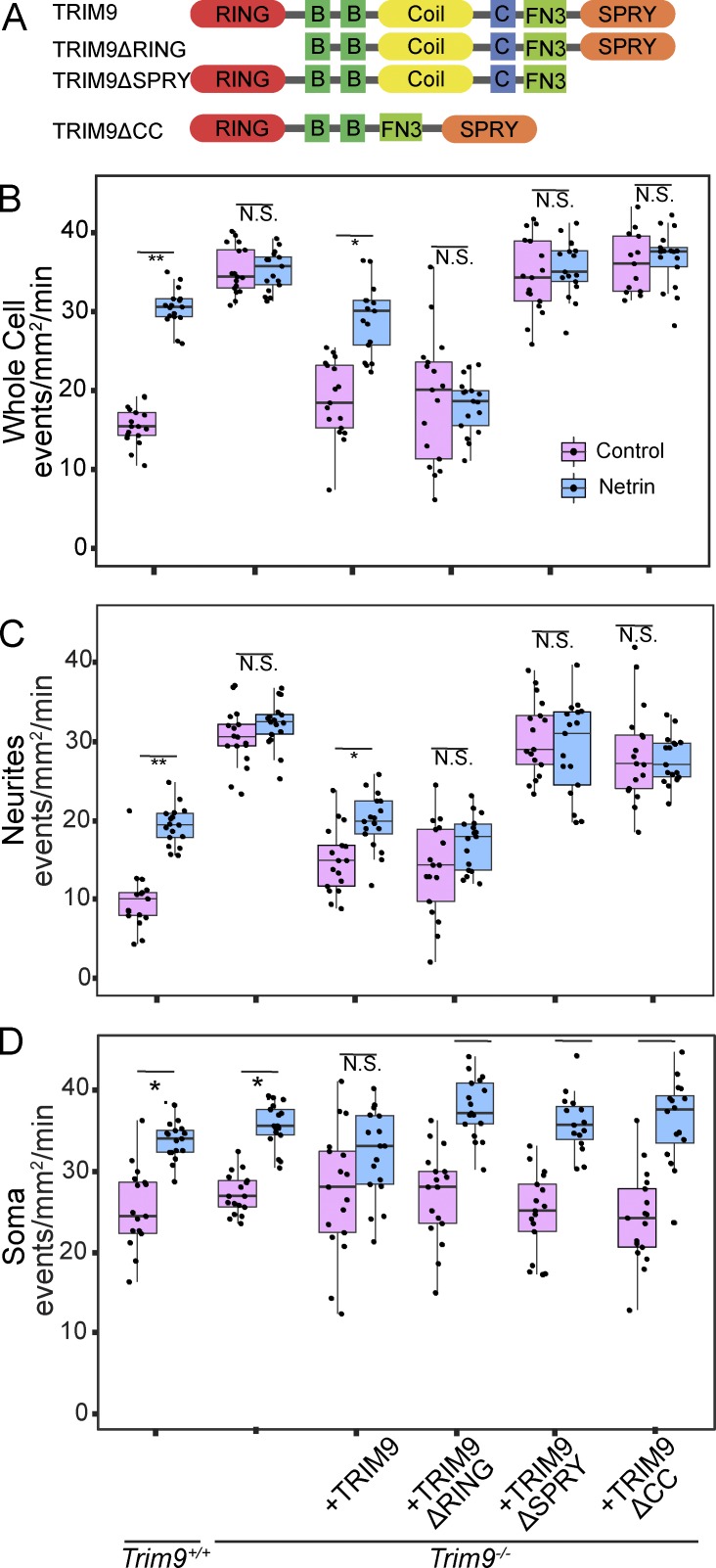
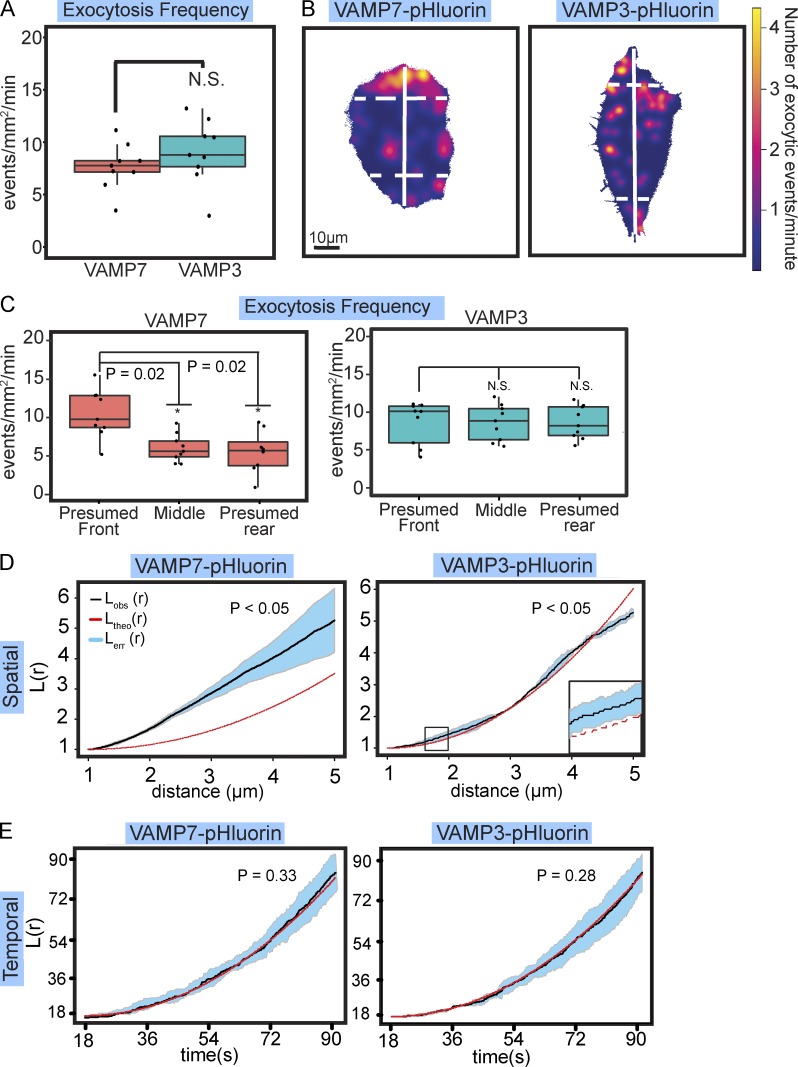
Neurite elongation and branching in developing neurons requires plasmalemma expansion, hypothesized to occur primarily via exocytosis. We posited that exocytosis in developing neurons and nonneuronal cells would exhibit distinct spatiotemporal organization. We exploited total internal reflection fluorescence microscopy to image vesicle-associated membrane protein (VAMP)-pHluorin-mediated exocytosis in mouse embryonic cortical neurons and interphase melanoma cells, and developed computer-vision software and statistical tools to uncover spatiotemporal aspects of exocytosis. Vesicle fusion behavior differed between vesicle types, cell types, developmental stages, and extracellular environments. Experiment-based mathematical calculations indicated that VAMP2-mediated vesicle fusion supplied excess material for the plasma membrane expansion that occurred early in neuronal morphogenesis, which was balanced by clathrin-mediated endocytosis. Spatial statistics uncovered distinct spatiotemporal regulation of exocytosis in the soma and neurites of developing neurons that was modulated by developmental stage, exposure to the guidance cue netrin-1, and the brain-enriched ubiquitin ligase tripartite motif 9. In melanoma cells, exocytosis occurred less frequently, with distinct spatial clustering patterns.
© 2018 Urbina et al.
Figures
References
-
- Alberts P., Rudge R., Irinopoulou T., Danglot L., Gauthier-Rouvière C., and Galli T.. 2006. Cdc42 and actin control polarized expression of TI-VAMP vesicles to neuronal growth cones and their fusion with the plasma membrane. Mol. Biol. Cell. 17:1194–1203. 10.1091/mbc.E05-07-0643 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
