

Eliminating Glutamatergic Input onto Horizontal Cells Changes the Dynamic Range and Receptive Field Organization of Mouse Retinal Ganglion Cells
- PMID: 29352045
- PMCID: PMC6705883
- DOI: 10.1523/JNEUROSCI.0141-17.2018
Eliminating Glutamatergic Input onto Horizontal Cells Changes the Dynamic Range and Receptive Field Organization of Mouse Retinal Ganglion Cells
Abstract
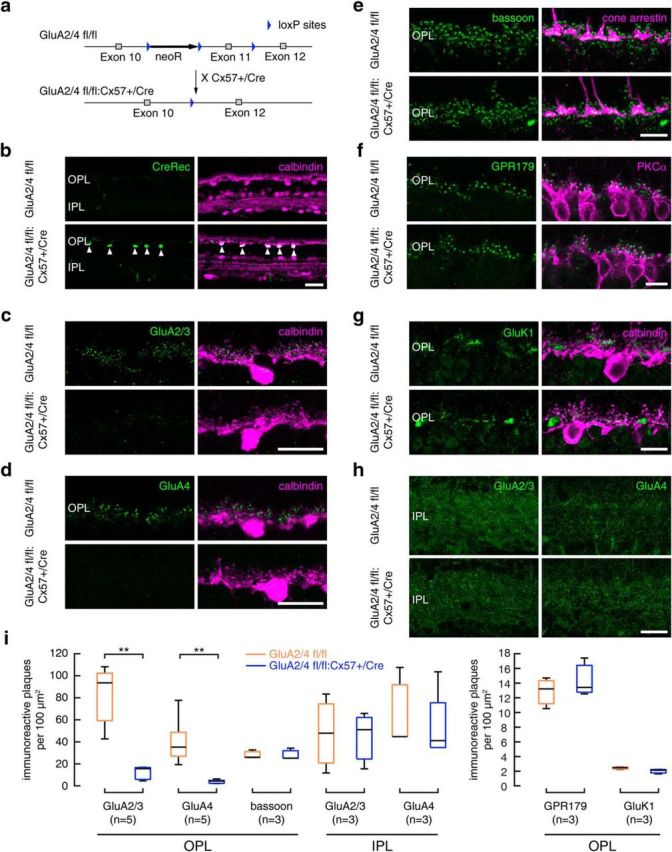
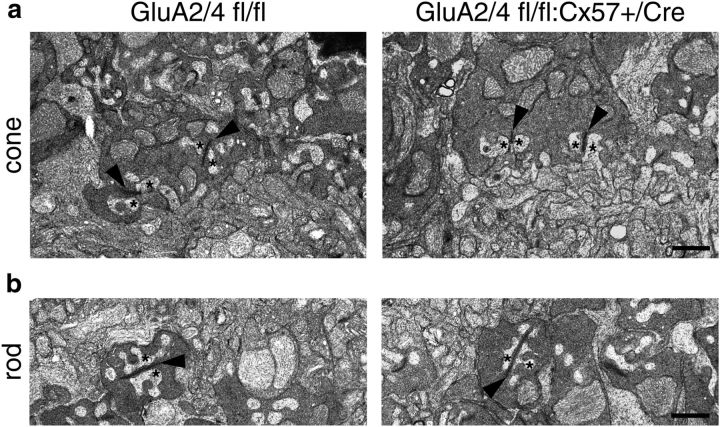
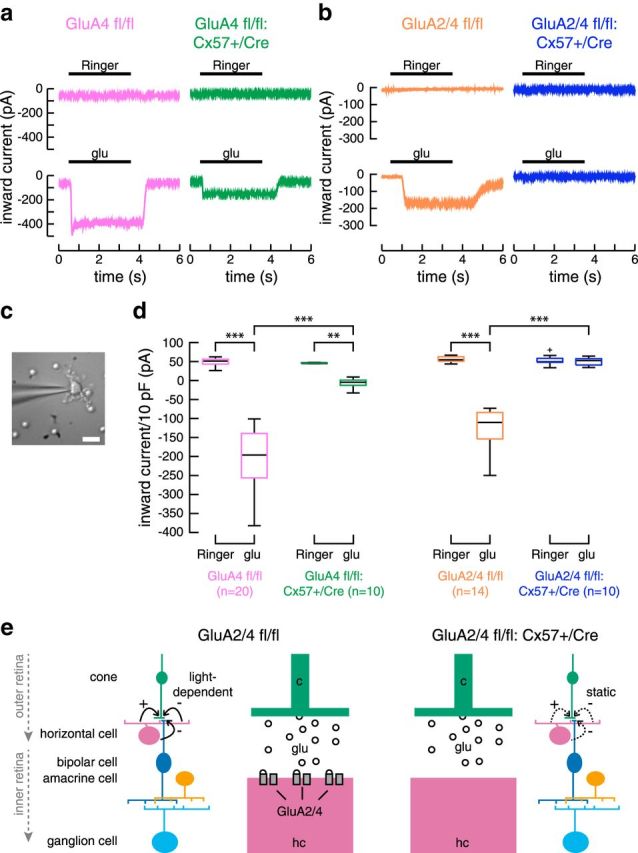
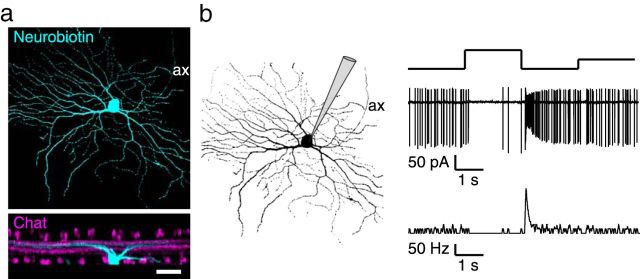
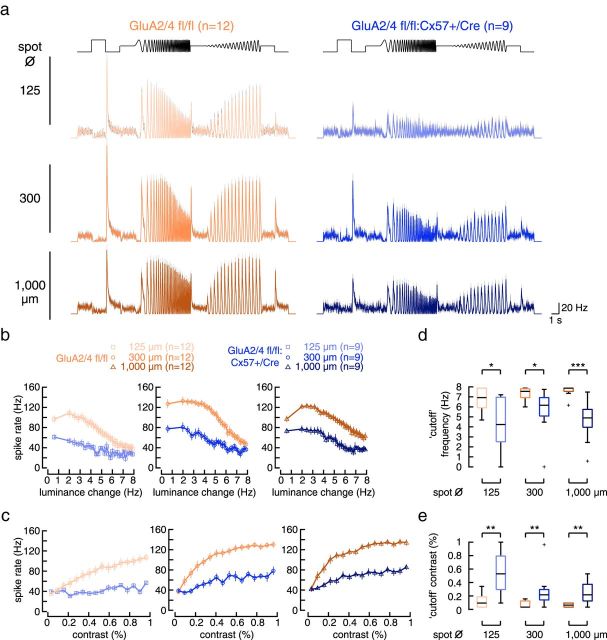
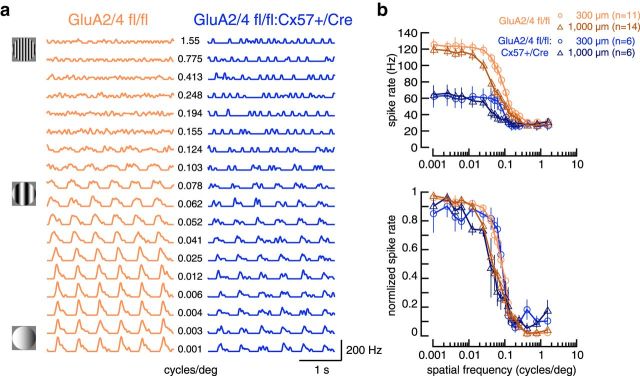
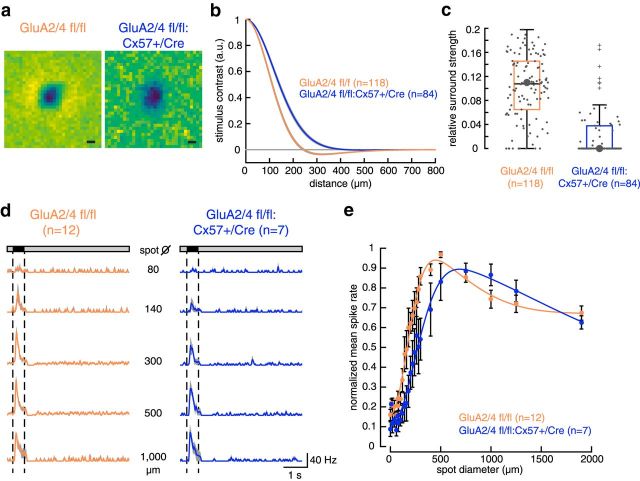
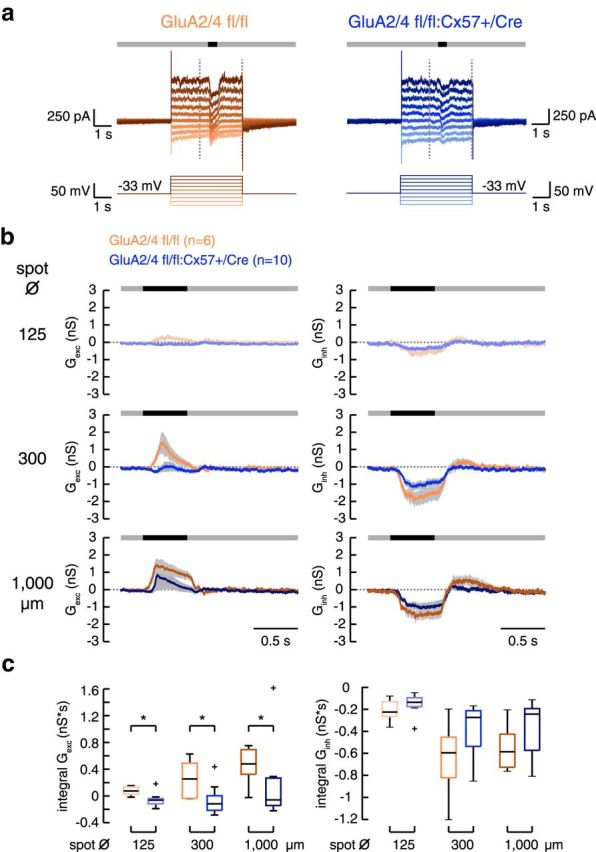
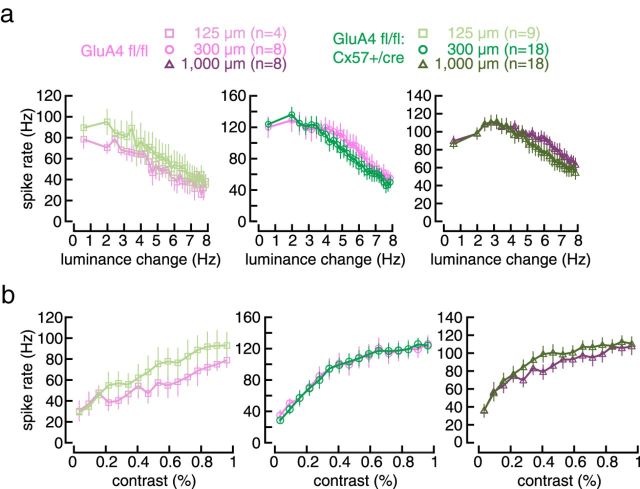
In the mammalian retina, horizontal cells receive glutamatergic inputs from many rod and cone photoreceptors and return feedback signals to them, thereby changing photoreceptor glutamate release in a light-dependent manner. Horizontal cells also provide feedforward signals to bipolar cells. It is unclear, however, how horizontal cell signals also affect the temporal, spatial, and contrast tuning in retinal output neurons, the ganglion cells. To study this, we generated a genetically modified mouse line in which we eliminated the light dependency of feedback by deleting glutamate receptors from mouse horizontal cells. This genetic modification allowed us to investigate the impact of horizontal cells on ganglion cell signaling independent of the actual mode of feedback in the outer retina and without pharmacological manipulation of signal transmission. In control and genetically modified mice (both sexes), we recorded the light responses of transient OFF-α retinal ganglion cells in the intact retina. Excitatory postsynaptic currents (EPSCs) were reduced and the cells were tuned to lower temporal frequencies and higher contrasts, presumably because photoreceptor output was attenuated. Moreover, receptive fields of recorded cells showed a significantly altered surround structure. Our data thus suggest that horizontal cells are responsible for adjusting the dynamic range of retinal ganglion cells and, together with amacrine cells, contribute to the center/surround organization of ganglion cell receptive fields in the mouse.SIGNIFICANCE STATEMENT Horizontal cells represent a major neuronal class in the mammalian retina and provide lateral feedback and feedforward signals to photoreceptors and bipolar cells, respectively. The mode of signal transmission remains controversial and, moreover, the contribution of horizontal cells to visual processing is still elusive. To address the question of how horizontal cells affect retinal output signals, we recorded the light responses of transient OFF-α retinal ganglion cells in a newly generated mouse line. In this mouse line, horizontal cell signals were no longer modulated by light. With light response recordings, we show that horizontal cells increase the dynamic range of retinal ganglion cells for contrast and temporal changes and contribute to the center/surround organization of their receptive fields.
Keywords: gain control; ganglion cells; glutamate receptor; horizontal cells; receptive field; retina.
Copyright © 2018 the authors 0270-6474/18/382015-14$15.00/0.
Figures
References
-
- Conover WJ, Iman RL (1981) Rank transformations as a bridge between parametric and nonparametric statistics. Am Stat 35:124–129. 10.1080/00031305.1981.10479327 - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases