

Mycolyltransferase from Mycobacterium tuberculosis in covalent complex with tetrahydrolipstatin provides insights into antigen 85 catalysis
- PMID: 29352107
- PMCID: PMC5846135
- DOI: 10.1074/jbc.RA117.001681
Mycolyltransferase from Mycobacterium tuberculosis in covalent complex with tetrahydrolipstatin provides insights into antigen 85 catalysis
Abstract
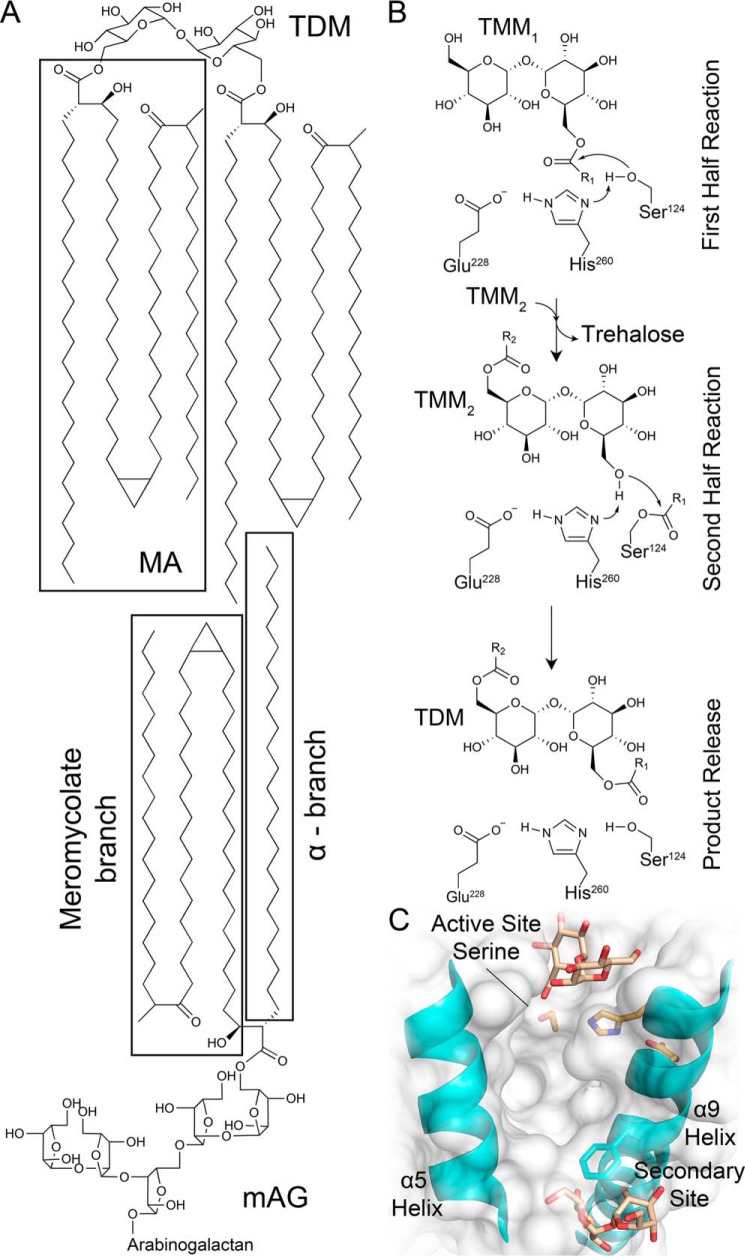
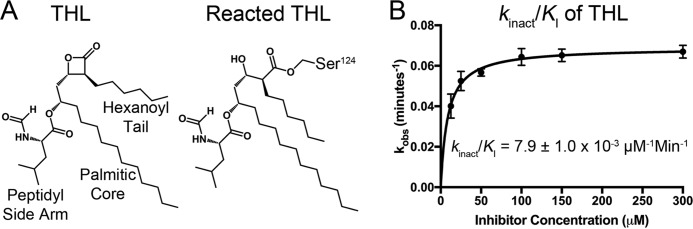
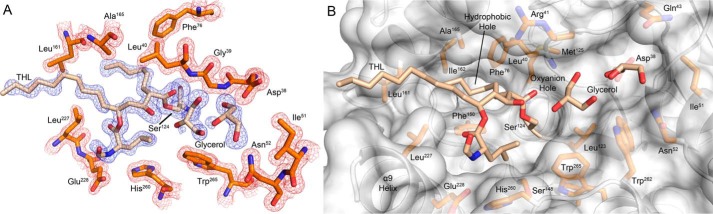
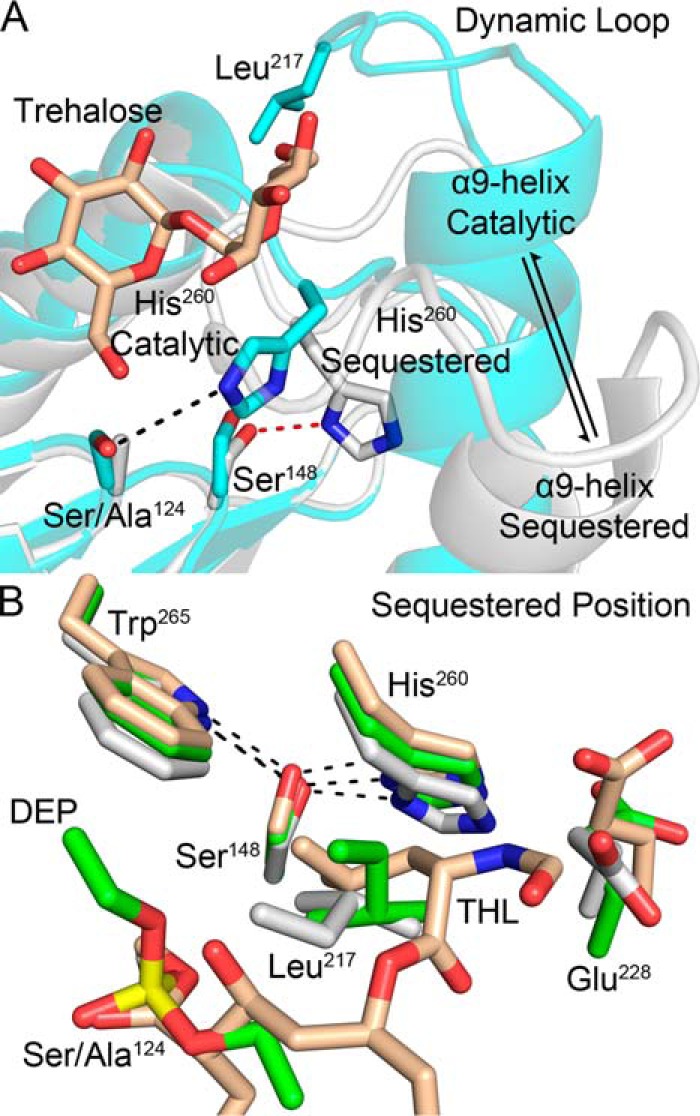
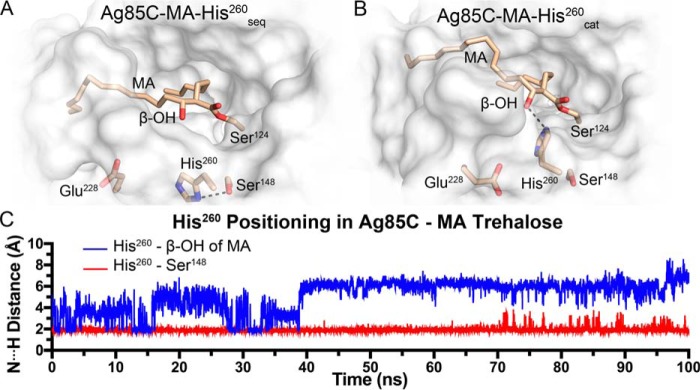
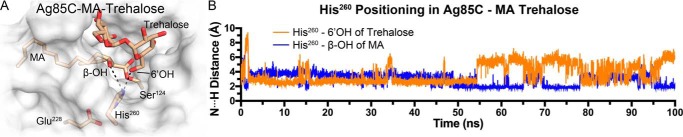
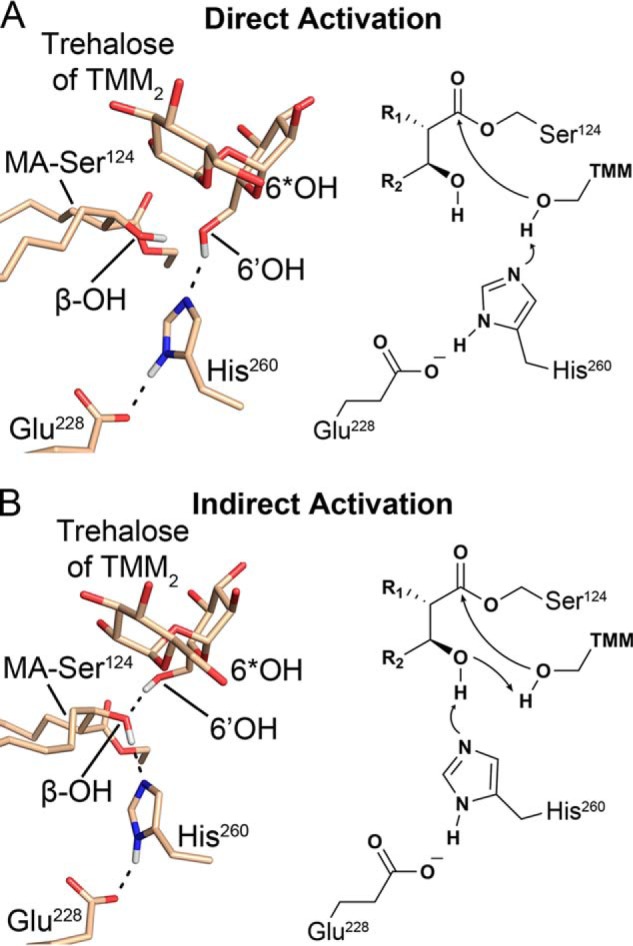
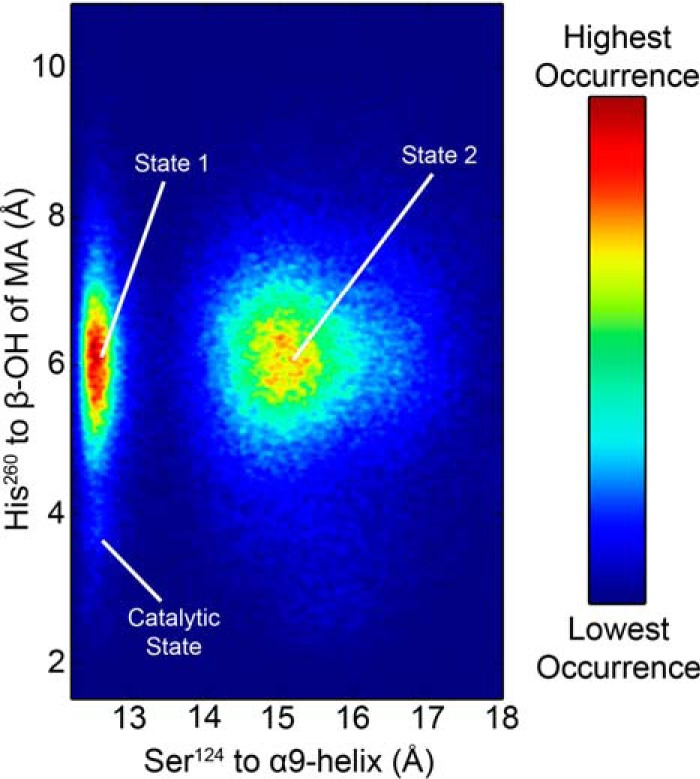
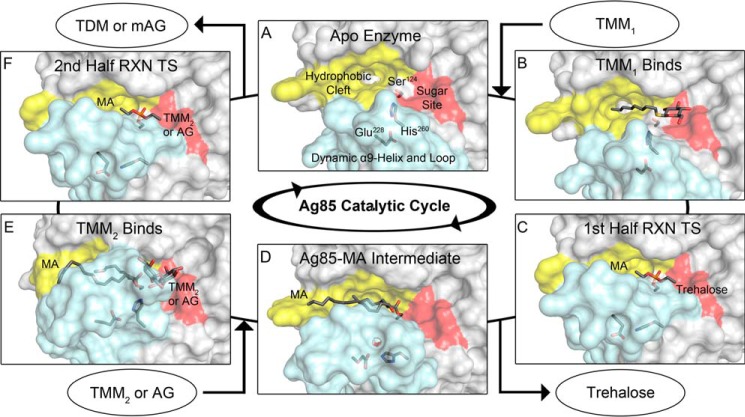
Mycobacterium tuberculosis antigen 85 (Ag85) enzymes catalyze the transfer of mycolic acid (MA) from trehalose monomycolate to produce the mycolyl arabinogalactan (mAG) or trehalose dimycolate (TDM). These lipids define the protective mycomembrane of mycobacteria. The current model of substrate binding within the active sites of Ag85s for the production of TDM is not sterically and geometrically feasible; additionally, this model does not account for the production of mAG. Furthermore, this model does not address how Ag85s limit the hydrolysis of the acyl-enzyme intermediate while catalyzing acyl transfer. To inform an updated model, we obtained an Ag85 acyl-enzyme intermediate structure that resembles the mycolated form. Here, we present a 1.45-Å X-ray crystal structure of M. tuberculosis Ag85C covalently modified by tetrahydrolipstatin (THL), an esterase inhibitor that suppresses M. tuberculosis growth and mimics structural attributes of MAs. The mode of covalent inhibition differs from that observed in the reversible inhibition of the human fatty-acid synthase by THL. Similarities between the Ag85-THL structure and previously determined Ag85C structures suggest that the enzyme undergoes structural changes upon acylation, and positioning of the peptidyl arm of THL limits hydrolysis of the acyl-enzyme adduct. Molecular dynamics simulations of the modeled mycolated-enzyme form corroborate the structural analysis. From these findings, we propose an alternative arrangement of substrates that rectifies issues with the previous model and suggest a direct role for the β-hydroxy of MA in the second half-reaction of Ag85 catalysis. This information affords the visualization of a complete mycolyltransferase catalytic cycle.
Keywords: acyltransferase; antigen 85; glycolipid; hydrolase; lipid esterase; mycolyltransferase; serine esterase; structural biology; tetrahydrolipstatin; α/β-hydrolase.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
