

Early impoverished environment delays the maturation of cerebral cortex
- PMID: 29352131
- PMCID: PMC5775315
- DOI: 10.1038/s41598-018-19459-y
Early impoverished environment delays the maturation of cerebral cortex
Erratum in
-
Author Correction: Early impoverished environment delays the maturation of cerebral cortex.Sci Rep. 2021 Dec 2;11(1):23532. doi: 10.1038/s41598-021-00562-6. Sci Rep. 2021. PMID: 34853339 Free PMC article. No abstract available.
Abstract
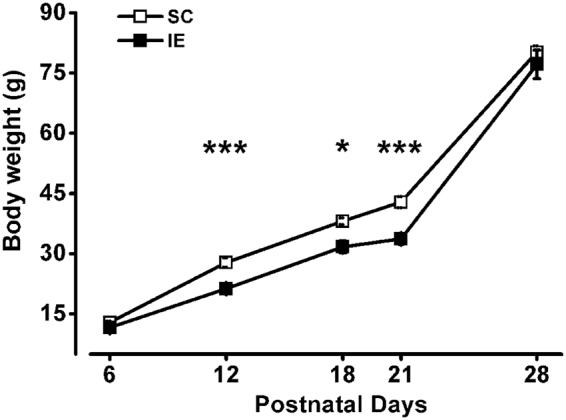
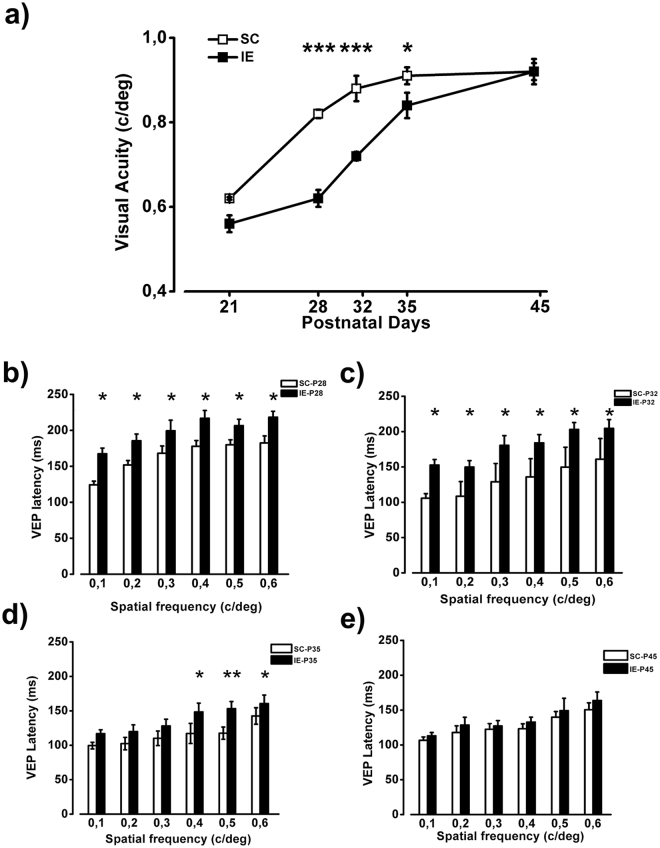
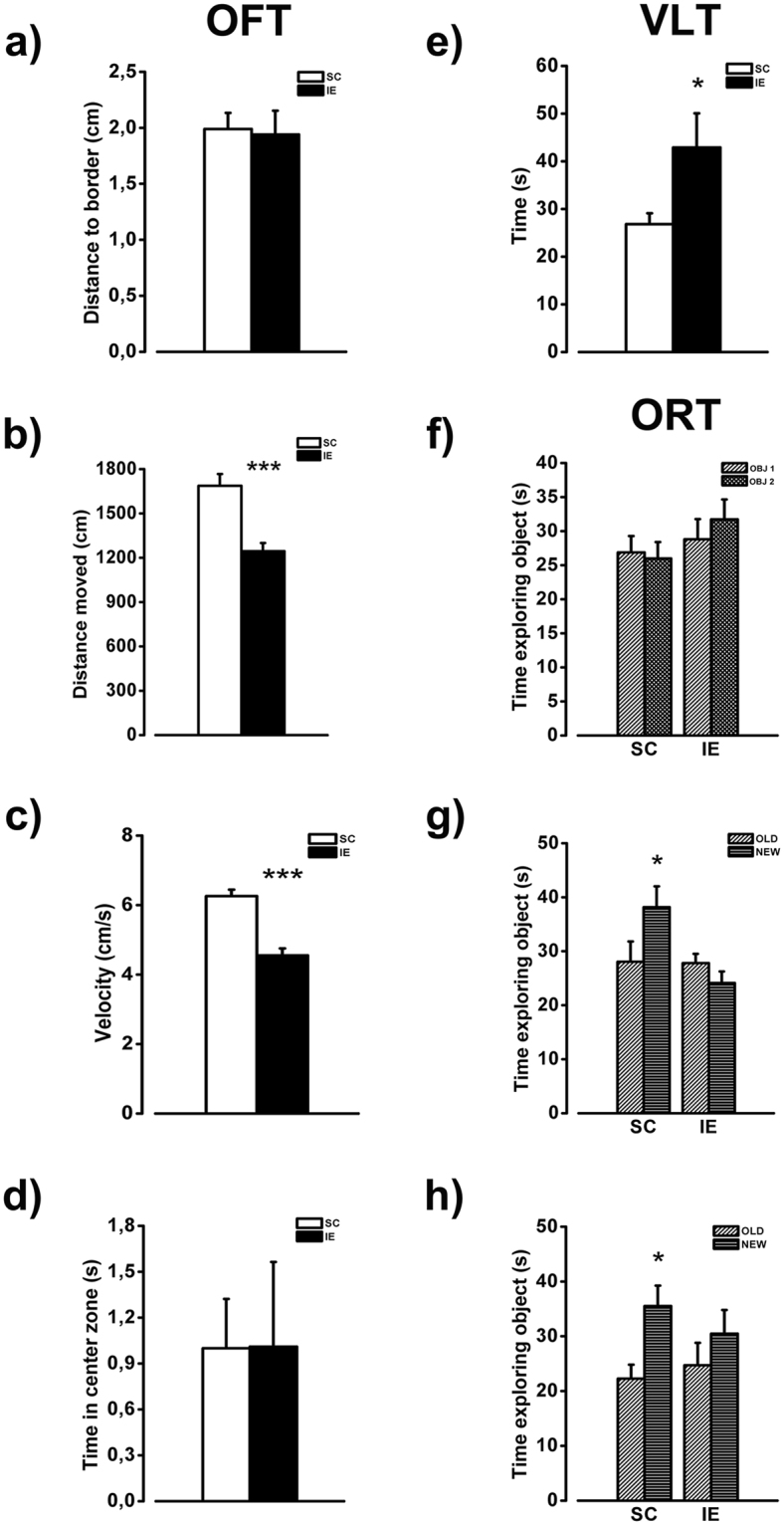
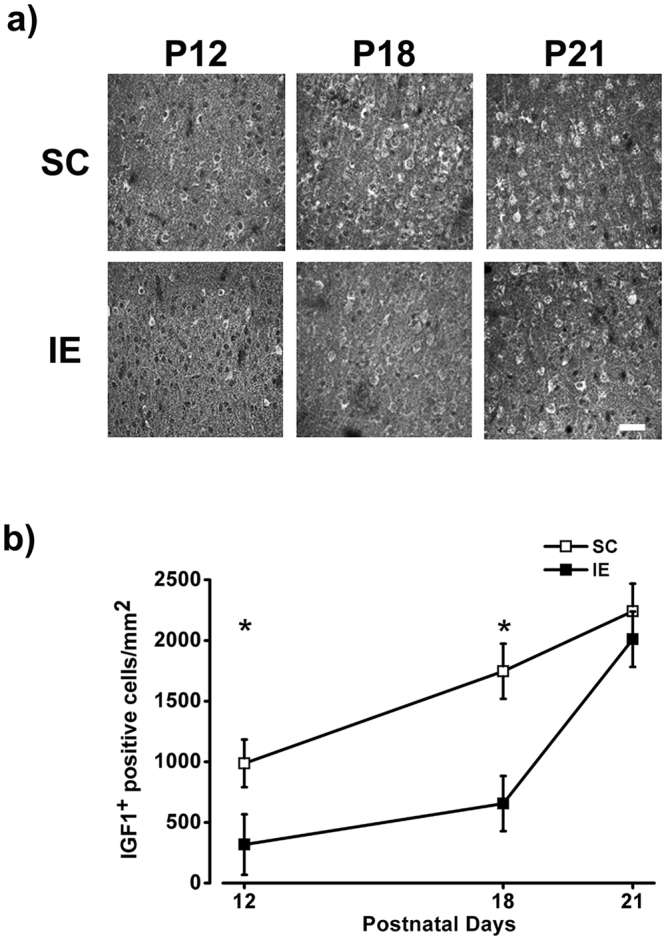
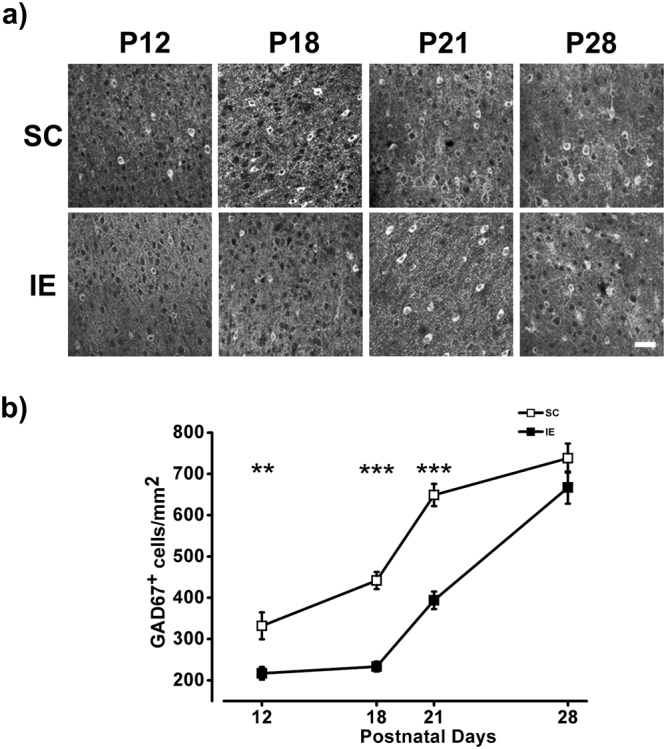
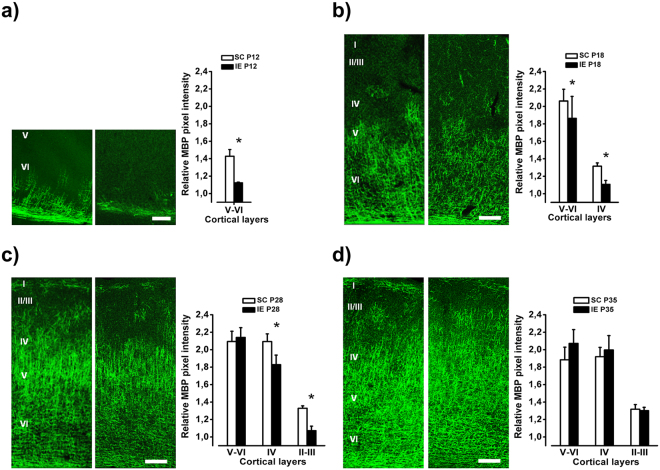
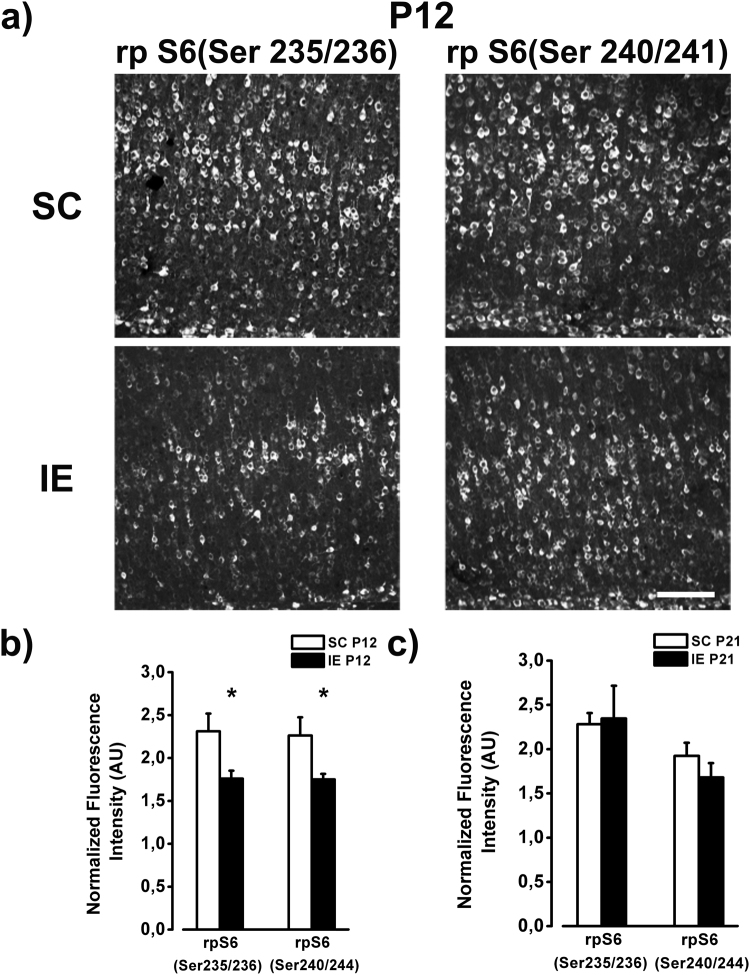
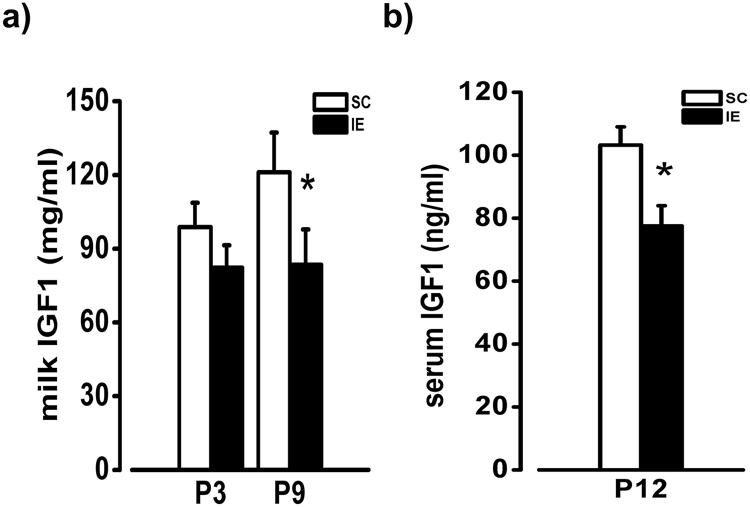
The influence of exposure to impoverished environments on brain development is unexplored since most studies investigated how environmental impoverishment affects adult brain. To shed light on the impact of early impoverishment on developmental trajectories of the nervous system, we developed a protocol of environmental impoverishment in which dams and pups lived from birth in a condition of reduced sensory-motor stimulation. Focusing on visual system, we measured two indexes of functional development, that is visual acuity, assessed by using Visual Evoked Potentials (VEPs), and VEP latency. In addition, we assessed in the visual cortex levels of Insulin-Like Growth Factor 1 (IGF-1) and myelin maturation, together with the expression of the GABA biosynthetic enzyme GAD67. We found that early impoverishment strongly delays visual acuity and VEP latency development. These functional changes were accompanied by a significant reduction of IGF-1 protein and GAD67 expression, as well as by delayed myelination of nerve fibers, in the visual cortex of impoverished pups. Thus, exposure to impoverished living conditions causes a significant alteration of developmental trajectories leading to a prominent delay of brain maturation. These results underscore the significance of adequate levels of environmental stimulation for the maturation of central nervous system.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Rosenzweig MR. Environmental complexity, cerebral change, and behavior. Am. Psychol. 1966;21:321–332. - PubMed
-
- Berardi N, Nicoletta B, Alessandro S, Lamberto M. Brain structural and functional development: genetics and experience. Developmental Medicine & Child Neurology. 2015;57:4–9. - PubMed
-
- Landi S, et al. Retinal functional development is sensitive to environmental enrichment: a role for BDNF. FASEB J. 2007;21:130–139. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
