

mRNA Translation Gone Awry: Translation Fidelity and Neurological Disease
- PMID: 29352613
- PMCID: PMC5834357
- DOI: 10.1016/j.tig.2017.12.007
mRNA Translation Gone Awry: Translation Fidelity and Neurological Disease
Abstract
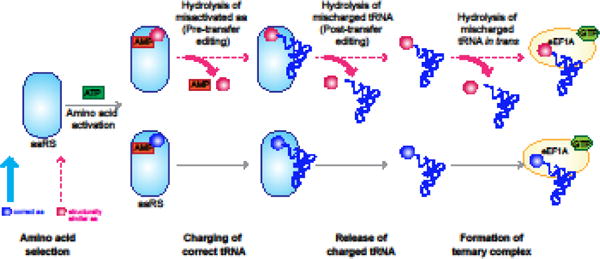
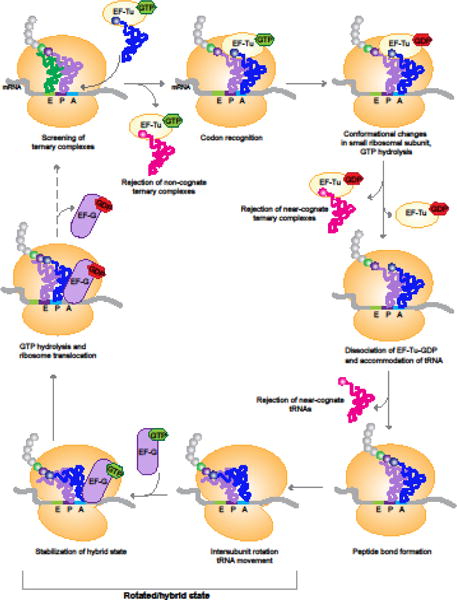
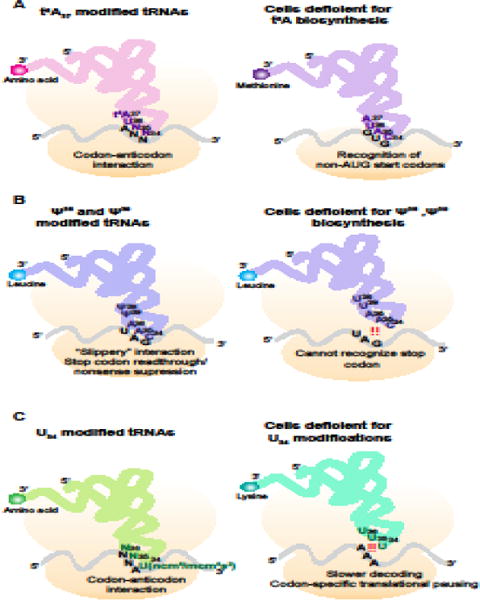
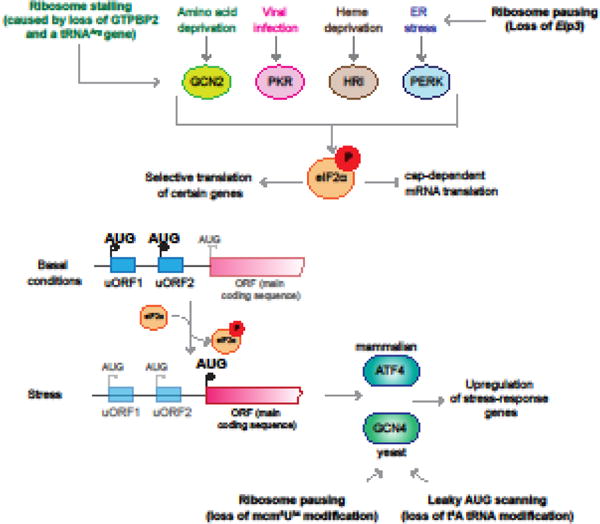
Errors during mRNA translation can lead to a reduction in the levels of functional proteins and an increase in deleterious molecules. Advances in next-generation sequencing have led to the discovery of rare genetic disorders, many caused by mutations in genes encoding the mRNA translation machinery, as well as to a better understanding of translational dynamics through ribosome profiling. We discuss here multiple neurological disorders that are linked to errors in tRNA aminoacylation and ribosome decoding. We draw on studies from genetic models, including yeast and mice, to enhance our understanding of the translational defects observed in these diseases. Finally, we emphasize the importance of tRNA, their associated enzymes, and the inextricable link between accuracy and efficiency in the maintenance of translational fidelity.
Keywords: elongation factor; mistranslation; neurodegeneration; ribosome stalling; tRNA modifications.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
References
-
- Rozov A, et al. New Structural Insights into Translational Miscoding. Trends Biochem Sci. 2016;41:798–814. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
