

The common bean COK-4 and the Arabidopsis FER kinase domain share similar functions in plant growth and defence
- PMID: 29352746
- PMCID: PMC6638044
- DOI: 10.1111/mpp.12659
The common bean COK-4 and the Arabidopsis FER kinase domain share similar functions in plant growth and defence
Abstract
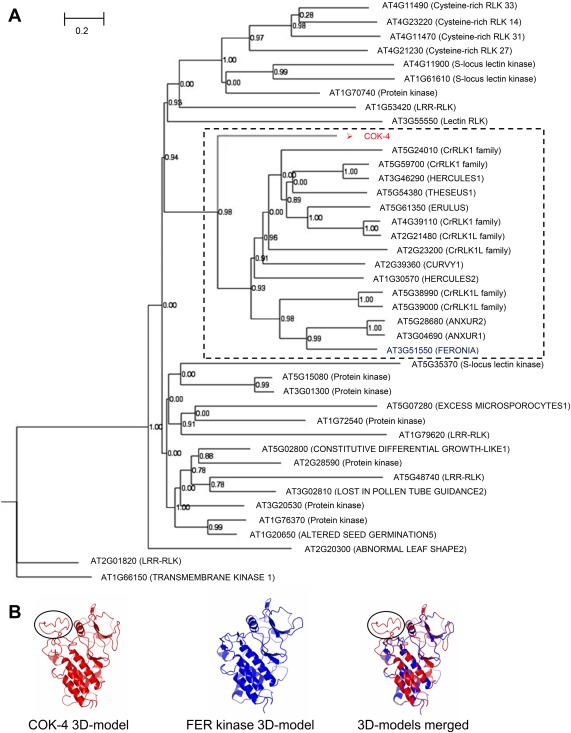
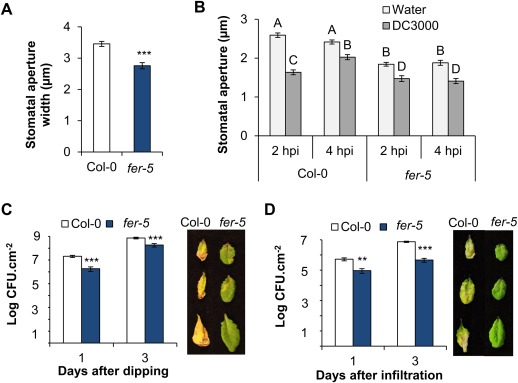
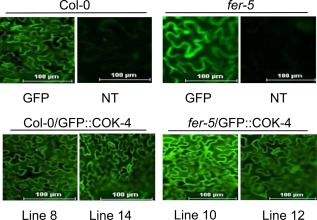
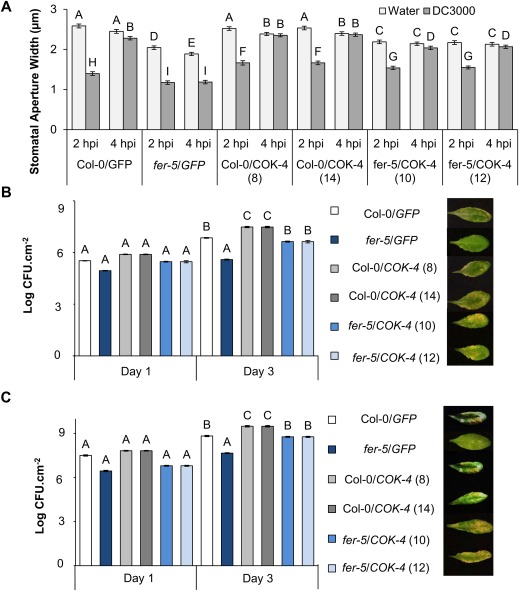
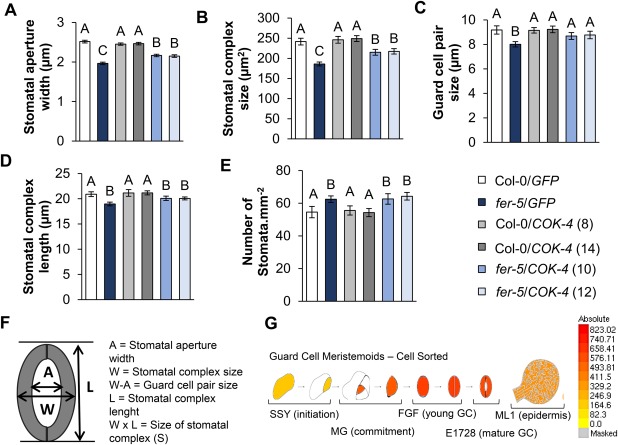
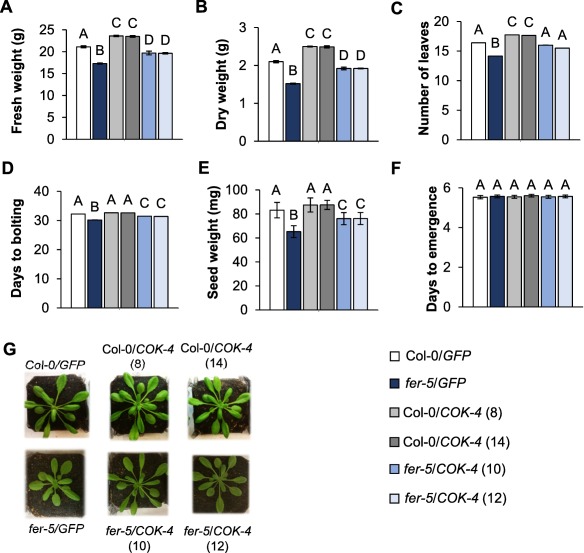
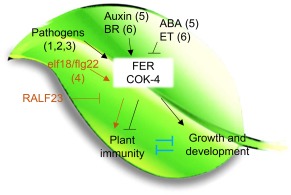
Receptor-like kinases are membrane proteins that can be shared by diverse signalling pathways. Among them, the Arabidopsis thaliana FERONIA (FER) plays a role in the balance between distinct signals to control growth and defence. We have found that COK-4, a putative kinase encoded in the common bean anthracnose resistance locus Co-4, which is transcriptionally regulated during the immune response, is highly similar to the kinase domain of FER. To assess whether COK-4 is a functional orthologue of FER, we expressed COK-4 in the wild-type Col-0 and the fer-5 mutant of Arabidopsis and evaluated FER-associated traits. We observed that fer-5 plants show an enhanced apoplastic and stomatal defence against Pseudomonas syringae. In addition, the fer-5 mutant shows reduced biomass, smaller guard cell size, greater number of stomata per leaf area, fewer leaves, faster transition to reproductive stage and lower seed weight per plant than the wild-type Col-0. Except for the stomatal complex length and number of stomata, COK-4 expression in fer-5 lines partially or completely rescued both defence and developmental defects of fer-5 to the wild-type level. Notably, COK-4 may have an additive effect to FER, as the expression of COK-4 in Col-0 resulted in enhanced defence and growth phenotypes in comparison with wild-type Col-0 plants. Altogether, these findings indicate that the common bean COK-4 shares at least some of the multiple functions of the Arabidopsis FER kinase domain, acting in both the induction of plant growth and regulation of plant defence.
Keywords: Arabidopsis thaliana; Co-4 locus; FERONIA; Phaseolus vulgaris; plant growth and development; plant immunity.
© 2018 BSPP AND JOHN WILEY & SONS LTD.
Figures
References
-
- Adrian, J. , Chang, J. , Ballenger, C.E. , Bargmann, B.O. , Alassimone, J. , Davies, K.A. , Lau, O.S. , Matos, J.L. , Hachez, C. , Lanctot, A. , Vatén, A. , Birnbaum, K.D. and Bergmann, D.C. (2015) Transcriptome dynamics of the stomatal lineage: birth, amplification, and termination of a self‐renewing population. Dev. Cell, 33, 107–118. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
