

Circadian- and Light-driven Metabolic Rhythms in Drosophila melanogaster
- PMID: 29355066
- PMCID: PMC6692290
- DOI: 10.1177/0748730417753003
Circadian- and Light-driven Metabolic Rhythms in Drosophila melanogaster
Abstract
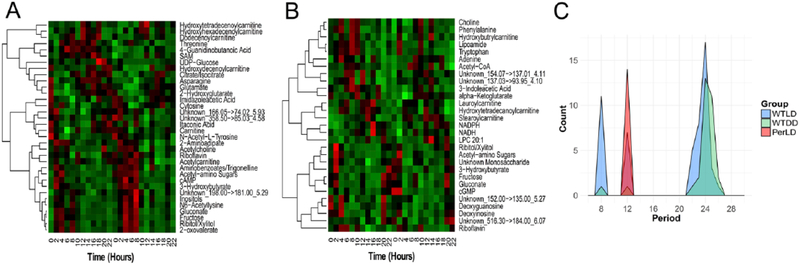
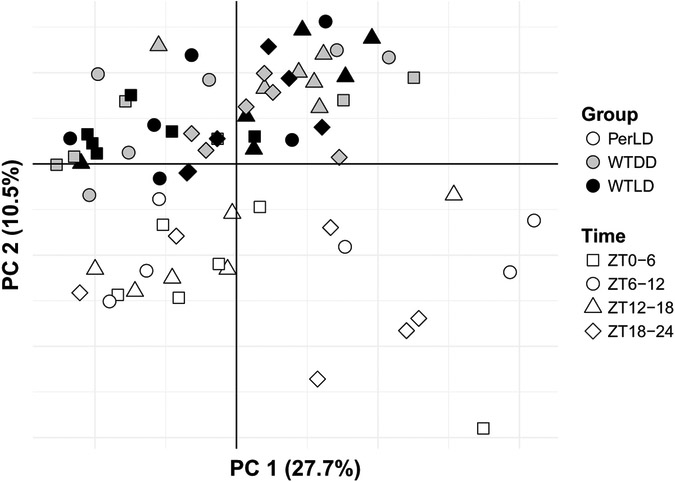
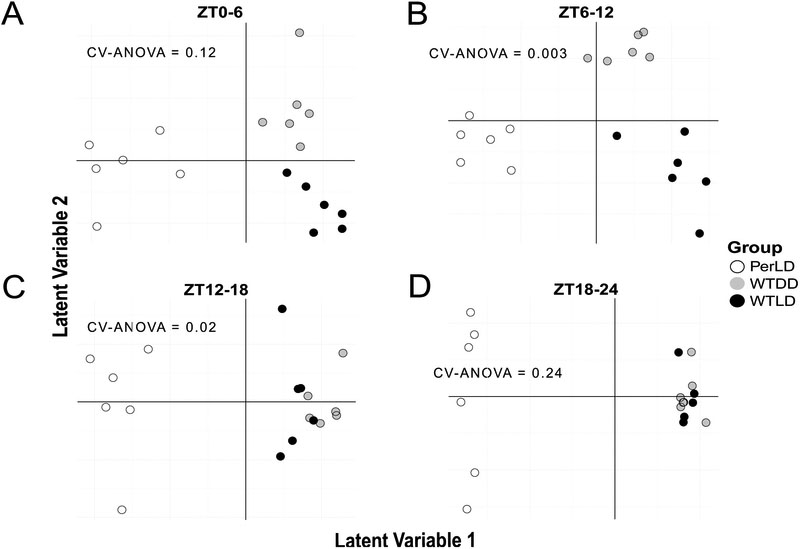
Complex interactions of environmental cues and transcriptional clocks drive rhythmicity in organismal physiology. Light directly affects the circadian clock; however, little is known about its relative role in controlling metabolic variations in vivo. Here we used high time-resolution sampling in Drosophila at every 2 h to measure metabolite outputs using a liquid-chromatography tandem mass spectrometry (LC-MS/MS) approach. Over 14% of detected metabolites oscillated with circadian periodicity under light-dark (LD) cycles. Many metabolites peaked shortly after lights-on, suggesting responsiveness to feeding and/or activity rather than the preactivity anticipation, as observed in previous transcriptomics analyses. Roughly 9% of measured metabolites uniquely oscillated under constant darkness (DD), suggesting that metabolite rhythms are associated with the transcriptional clock machinery. Strikingly, metabolome differences between LD and constant darkness were observed only during the light phase, highlighting the importance of photic input. Clock mutant flies exhibited strong 12-h ultradian rhythms, including 4 carbohydrate species with circadian periods in wild-type flies, but lacked 24-h circadian metabolic oscillations. A meta-analysis of these results with previous circadian metabolomics experiments uncovered the possibility of conserved rhythms in amino acids, keto-acids, and sugars across flies, mice, and humans and provides a basis for exploring the chrono-metabolic connection with powerful genetic tools in Drosophila.
Keywords: Drosophila; LC-MS metabolomics; circadian clock; conserved oscillators; photic metabolism; ultradian rhythms.
Conflict of interest statement
CONFLICT OF INTEREST STATEMENT
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Figures


References
-
- Agudelo LZ, Femenia T, Orhan F, Porsmyr-Palmertz M, Goiny M, Martinez-Redondo V, Correia JC, Izadi M, Bhat M, Schuppe-Koistinen I, et al. (2014) Skeletal muscle PGC-lαl modulates kynurenine metabolism and mediates resilience to stress-induced depression. Cell 159:33–45. - PubMed
-
- Ang JE, Revell V, Mann A, Mäntele S, Otway DT, Johnston JD, Thumser AE, Skene DJ, and Raynaud F (2012) Identification of human plasma metabolites exhibiting time-of-day variation using an untargeted liquid chromatography-mass spectrometry metabolomic approach. Chronobiol Int 29:868–881. - PMC - PubMed
-
- Bass J (2012) Circadian topology of metabolism. Nature 491:348–356. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
