

Regional and subtype-dependent miRNA signatures in sporadic Creutzfeldt-Jakob disease are accompanied by alterations in miRNA silencing machinery and biogenesis
- PMID: 29357384
- PMCID: PMC5794191
- DOI: 10.1371/journal.ppat.1006802
Regional and subtype-dependent miRNA signatures in sporadic Creutzfeldt-Jakob disease are accompanied by alterations in miRNA silencing machinery and biogenesis
Abstract
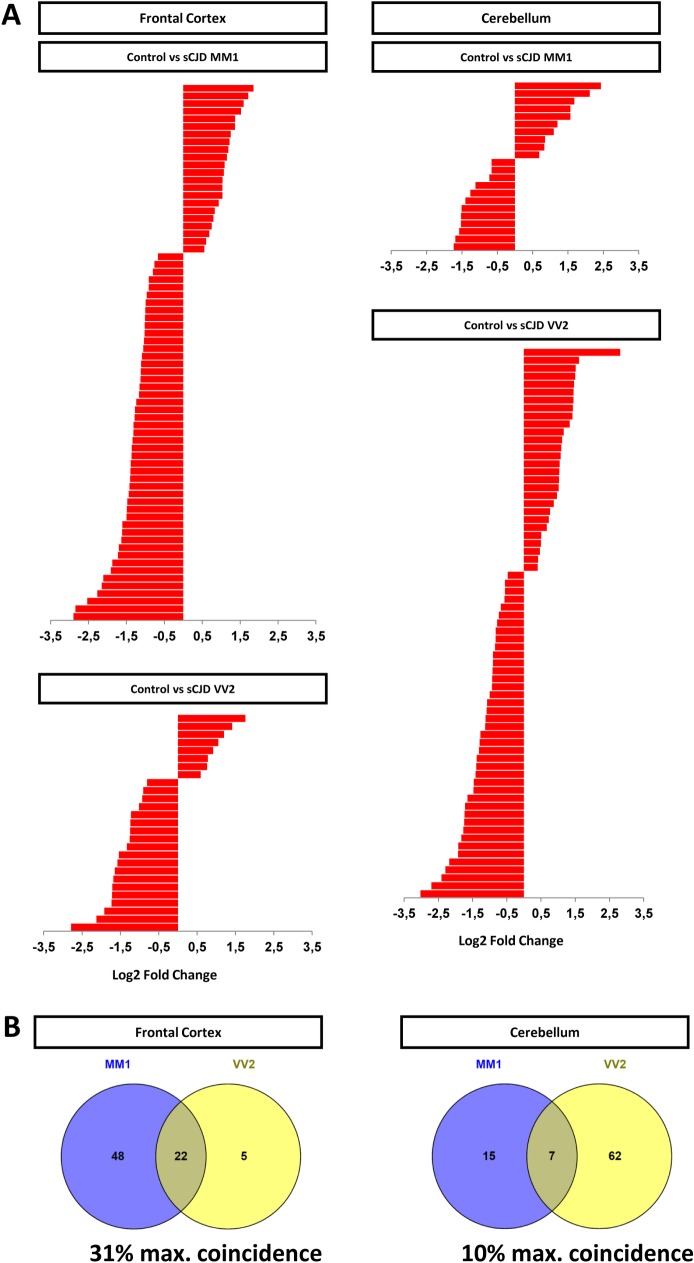
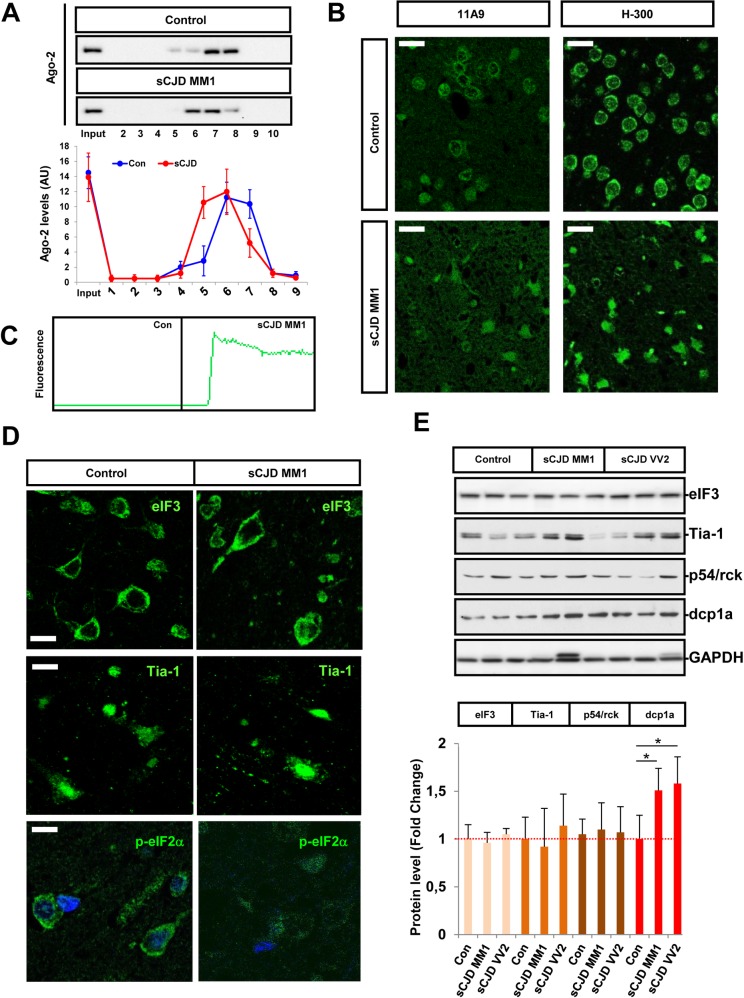
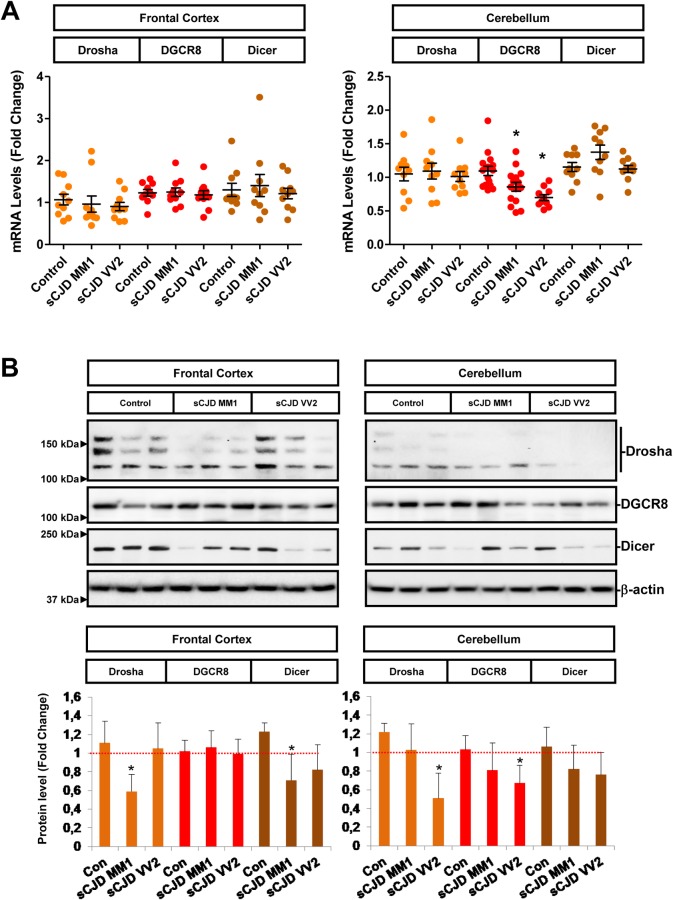
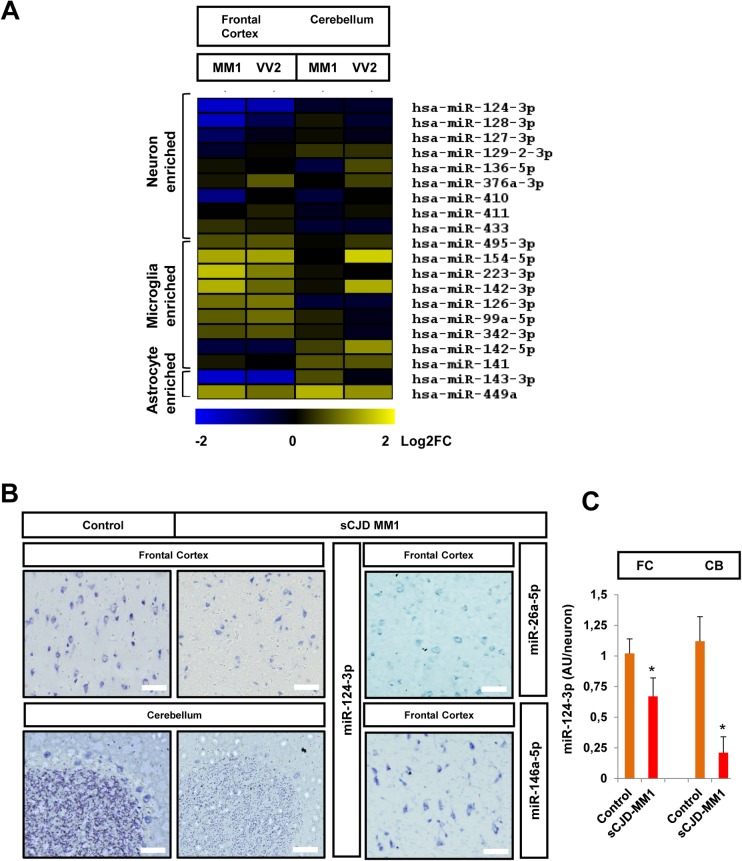
Increasing evidence indicates that microRNAs (miRNAs) are contributing factors to neurodegeneration. Alterations in miRNA signatures have been reported in several neurodegenerative dementias, but data in prion diseases are restricted to ex vivo and animal models. The present study identified significant miRNA expression pattern alterations in the frontal cortex and cerebellum of sporadic Creutzfeldt-Jakob disease (sCJD) patients. These changes display a highly regional and disease subtype-dependent regulation that correlates with brain pathology. We demonstrate that selected miRNAs are enriched in sCJD isolated Argonaute(Ago)-binding complexes in disease, indicating their incorporation into RNA-induced silencing complexes, and further suggesting their contribution to disease-associated gene expression changes. Alterations in the miRNA-mRNA regulatory machinery and perturbed levels of miRNA biogenesis key components in sCJD brain samples reported here further implicate miRNAs in sCJD gene expression (de)regulation. We also show that a subset of sCJD-altered miRNAs are commonly changed in Alzheimer's disease, dementia with Lewy bodies and fatal familial insomnia, suggesting potential common mechanisms underlying these neurodegenerative processes. Additionally, we report no correlation between brain and cerebrospinal fluid (CSF) miRNA-profiles in sCJD, indicating that CSF-miRNA profiles do not faithfully mirror miRNA alterations detected in brain tissue of human prion diseases. Finally, utilizing a sCJD MM1 mouse model, we analyzed the miRNA deregulation patterns observed in sCJD in a temporal manner. While fourteen sCJD-related miRNAs were validated at clinical stages, only two of those were changed at early symptomatic phase, suggesting that the miRNAs altered in sCJD may contribute to later pathogenic processes. Altogether, the present work identifies alterations in the miRNA network, biogenesis and miRNA-mRNA silencing machinery in sCJD, whereby contributions to disease mechanisms deserve further investigation.
Conflict of interest statement
The authors have declared that no competing interests exist."
Figures

Similar articles
-
Subtype and regional regulation of prion biomarkers in sporadic Creutzfeldt-Jakob disease.Neuropathol Appl Neurobiol. 2015 Aug;41(5):631-45. doi: 10.1111/nan.12175. Epub 2015 Apr 30. Neuropathol Appl Neurobiol. 2015. PMID: 25134744
-
PrP mRNA and protein expression in brain and PrP(c) in CSF in Creutzfeldt-Jakob disease MM1 and VV2.Prion. 2013 Sep-Oct;7(5):383-93. doi: 10.4161/pri.26416. Epub 2013 Sep 18. Prion. 2013. PMID: 24047819 Free PMC article.
-
RNA editing alterations define manifestation of prion diseases.Proc Natl Acad Sci U S A. 2019 Sep 24;116(39):19727-19735. doi: 10.1073/pnas.1803521116. Epub 2019 Sep 6. Proc Natl Acad Sci U S A. 2019. PMID: 31492812 Free PMC article.
-
Clinical findings and diagnostic tests in Creutzfeldt-Jakob disease and variant Creutzfeldt-Jakob disease.Folia Neuropathol. 2004;42 Suppl B:24-38. Folia Neuropathol. 2004. PMID: 16903140 Review.
-
Genetic and Transcriptomic Profiles of Inflammation in Neurodegenerative Diseases: Alzheimer, Parkinson, Creutzfeldt-Jakob and Tauopathies.Int J Mol Sci. 2016 Feb 4;17(2):206. doi: 10.3390/ijms17020206. Int J Mol Sci. 2016. PMID: 26861289 Free PMC article. Review.
Cited by
-
MicroRNA Alterations in the Brain and Body Fluids of Humans and Animal Prion Disease Models: Current Status and Perspectives.Front Aging Neurosci. 2018 Jul 23;10:220. doi: 10.3389/fnagi.2018.00220. eCollection 2018. Front Aging Neurosci. 2018. PMID: 30083102 Free PMC article.
-
MicroRNAs in Cardiac Autophagy: Small Molecules and Big Role.Cells. 2018 Aug 11;7(8):104. doi: 10.3390/cells7080104. Cells. 2018. PMID: 30103495 Free PMC article. Review.
-
Identification of circulating microRNA signatures as potential biomarkers in the serum of elk infected with chronic wasting disease.Sci Rep. 2019 Dec 23;9(1):19705. doi: 10.1038/s41598-019-56249-6. Sci Rep. 2019. PMID: 31873177 Free PMC article.
-
Epigenomic Modifications in Modern and Ancient Genomes.Genes (Basel). 2022 Jan 20;13(2):178. doi: 10.3390/genes13020178. Genes (Basel). 2022. PMID: 35205223 Free PMC article. Review.
-
Exploring the Relationship Between Sporadic Creutzfeldt-Jakob Disease and Gut Microbiota Through a Mendelian Randomization Study.Mol Neurobiol. 2025 Feb;62(2):1945-1959. doi: 10.1007/s12035-024-04376-1. Epub 2024 Jul 25. Mol Neurobiol. 2025. PMID: 39052184
References
-
- Puoti G, Bizzi A, Forloni G, Safar JG, Tagliavini F, Gambetti P. Sporadic human prion diseases: Molecular insights and diagnosis. Lancet Neurol. 2012;11: 618–628. doi: 10.1016/S1474-4422(12)70063-7 - DOI - PubMed
-
- Gambetti P, Kong Q, Zou W, Parchi P, Chen SG. Sporadic and familial CJD: Classification and characterisation. British Medical Bulletin. 2003. pp. 213–239. doi: 10.1093/bmb/66.1.213 - DOI - PubMed
-
- Parchi P, De Boni L, Saverioni D, Cohen ML, Ferrer I, Gambetti P, et al. Consensus classification of human prion disease histotypes allows reliable identification of molecular subtypes: An inter-rater study among surveillance centres in Europe and USA. Acta Neuropathol. 2012;124: 517–529. doi: 10.1007/s00401-012-1002-8 - DOI - PMC - PubMed
-
- Parchi P, Giese a, Capellari S, Brown P, Schulz-Schaeffer W, Windl O, et al. Classification of sporadic Creutzfeldt-Jakob disease based on molecular and phenotypic analysis of 300 subjects. Ann Neurol. 1999;46: 224–233. doi: 10.1002/1531-8249(199908)46:2<224::AID-ANA12>3.0.CO;2-W - DOI - PubMed
-
- Llorens F, Lopez-Gonzalez I, Thune K, Carmona M, Zafar S, Andeoletti O, et al. Subtype and regional-specific neuroinflammation in sporadic creutzfeldt-jakob disease. Front Aging Neurosci. 2014;6 doi: 10.3389/fnagi.2014.00198 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
