

Non-classical Immunity Controls Microbiota Impact on Skin Immunity and Tissue Repair
- PMID: 29358051
- PMCID: PMC6034182
- DOI: 10.1016/j.cell.2017.12.033
Non-classical Immunity Controls Microbiota Impact on Skin Immunity and Tissue Repair
Abstract
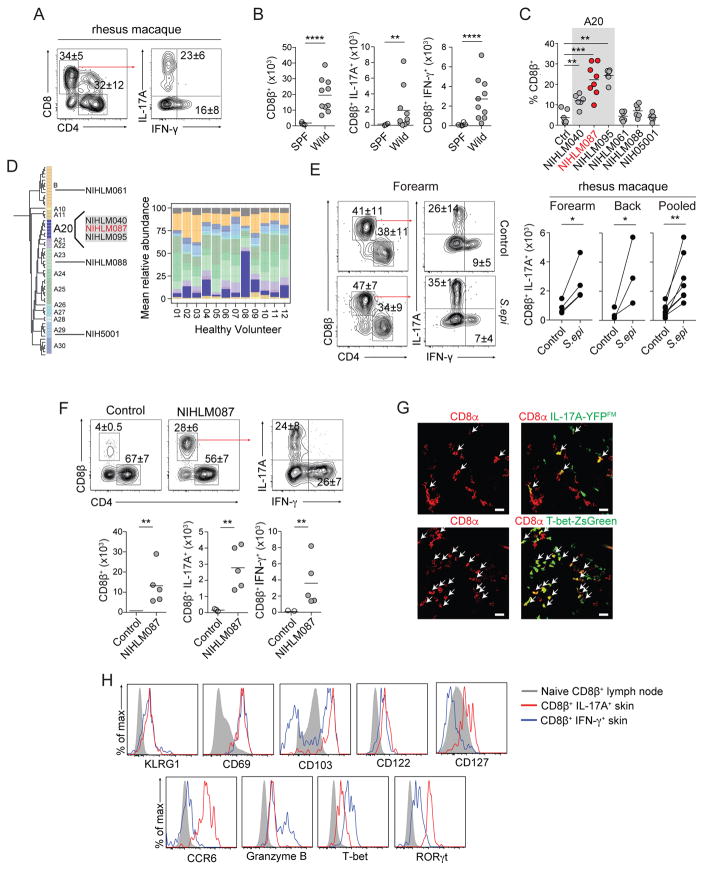
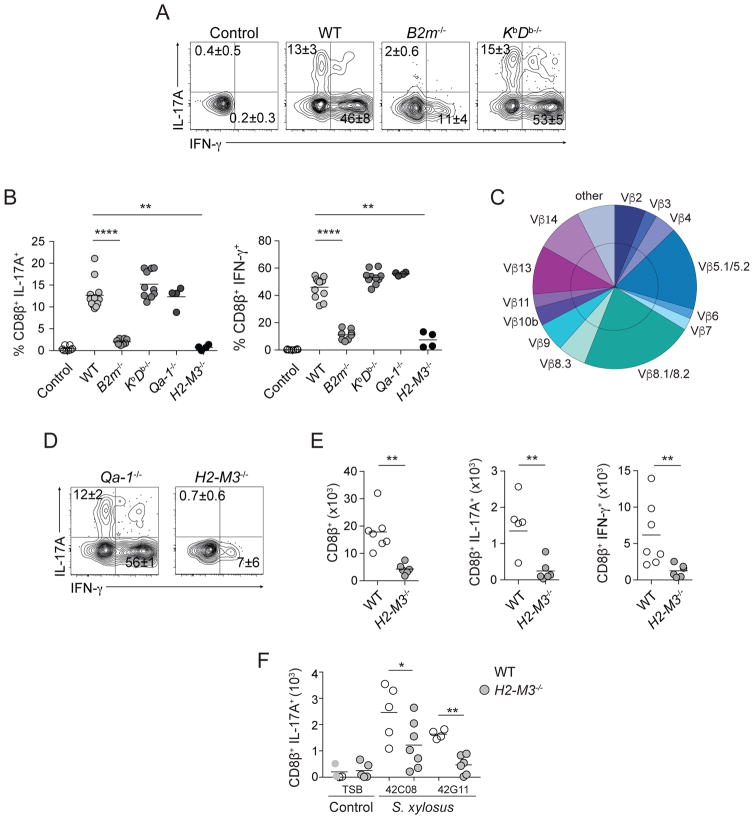
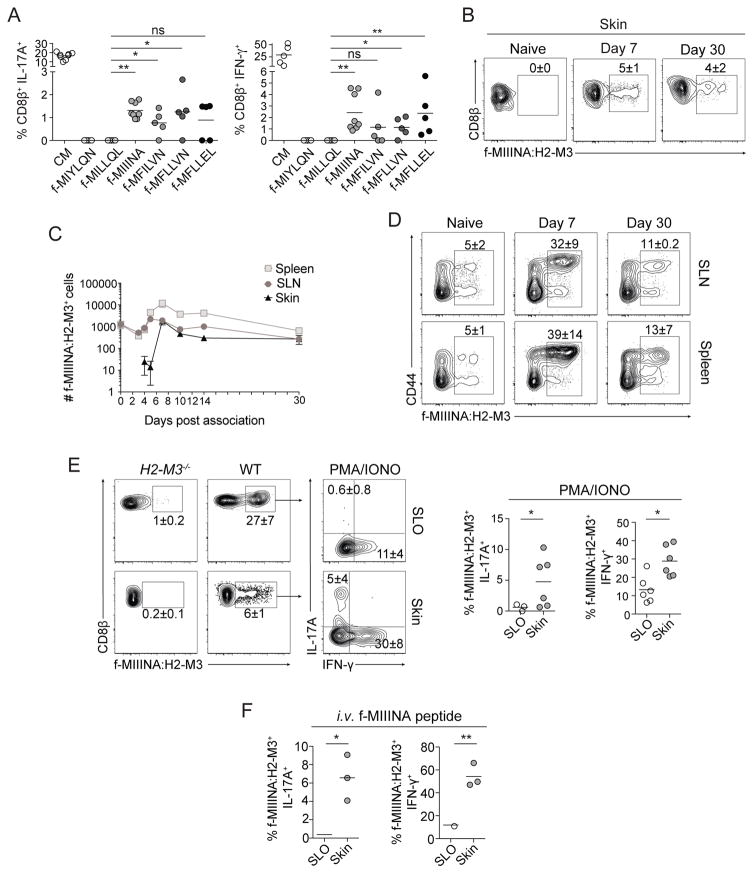
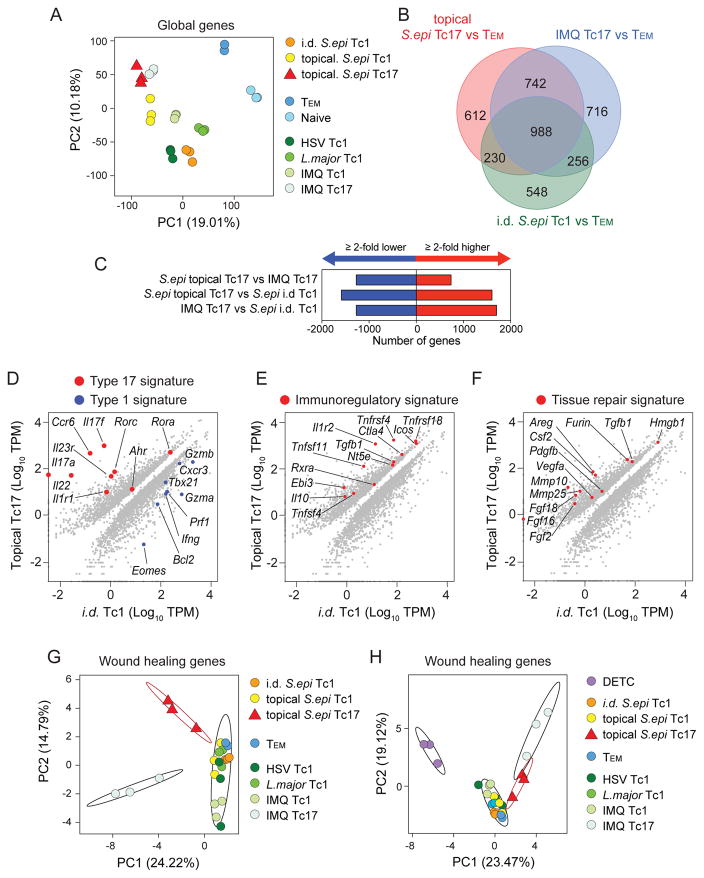
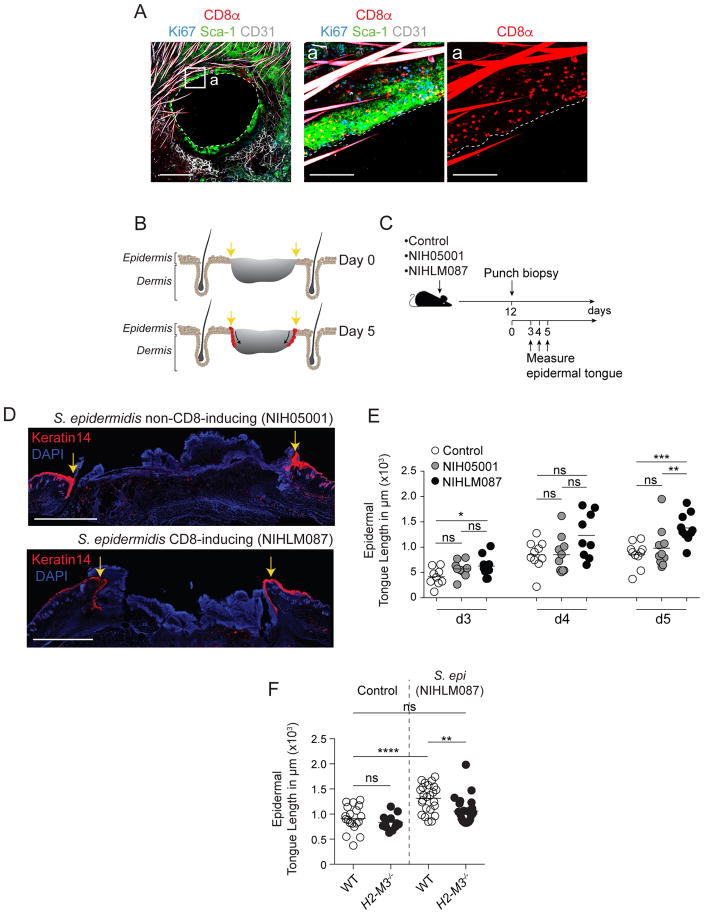
Mammalian barrier surfaces are constitutively colonized by numerous microorganisms. We explored how the microbiota was sensed by the immune system and the defining properties of such responses. Here, we show that a skin commensal can induce T cell responses in a manner that is restricted to non-classical MHC class I molecules. These responses are uncoupled from inflammation and highly distinct from pathogen-induced cells. Commensal-specific T cells express a defined gene signature that is characterized by expression of effector genes together with immunoregulatory and tissue-repair signatures. As such, non-classical MHCI-restricted commensal-specific immune responses not only promoted protection to pathogens, but also accelerated skin wound closure. Thus, the microbiota can induce a highly physiological and pleiotropic form of adaptive immunity that couples antimicrobial function with tissue repair. Our work also reveals that non-classical MHC class I molecules, an evolutionarily ancient arm of the immune system, can promote homeostatic immunity to the microbiota.
Keywords: H2-M3; MHCIb; Staphylococcus epidermidis; microbiota; non-classical MHC class I; skin immunity; tissue repair.
Published by Elsevier Inc.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
Skin Commensal Antigens: Taking the Road Less Traveled.Trends Immunol. 2018 Apr;39(4):259-261. doi: 10.1016/j.it.2018.02.001. Epub 2018 Feb 22. Trends Immunol. 2018. PMID: 29478772 Free PMC article.
-
Killer T cells show their kinder side.Nature. 2018 Mar 29;555(7698):594-595. doi: 10.1038/d41586-018-03510-z. Nature. 2018. PMID: 29595806 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
