

Lipids Shape the Electron Acceptor-Binding Site of the Peripheral Membrane Protein Dihydroorotate Dehydrogenase
- PMID: 29358052
- PMCID: PMC5856493
- DOI: 10.1016/j.chembiol.2017.12.012
Lipids Shape the Electron Acceptor-Binding Site of the Peripheral Membrane Protein Dihydroorotate Dehydrogenase
Abstract
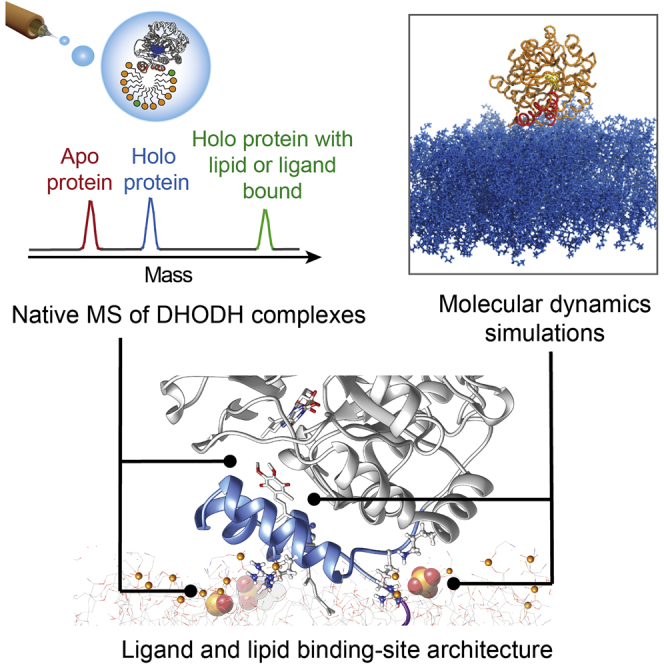
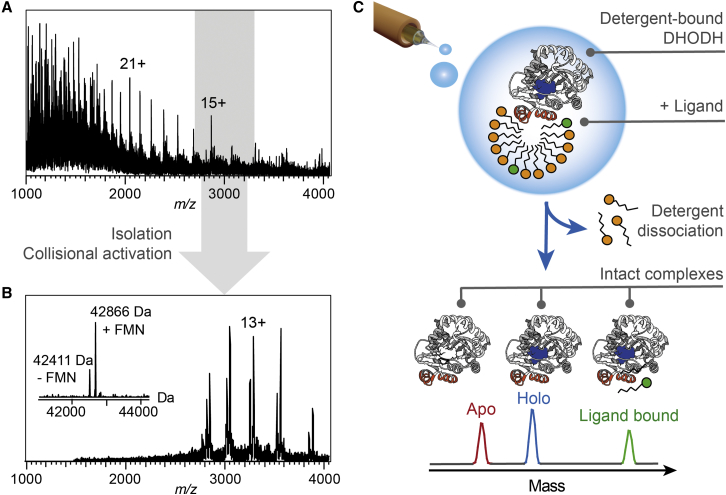
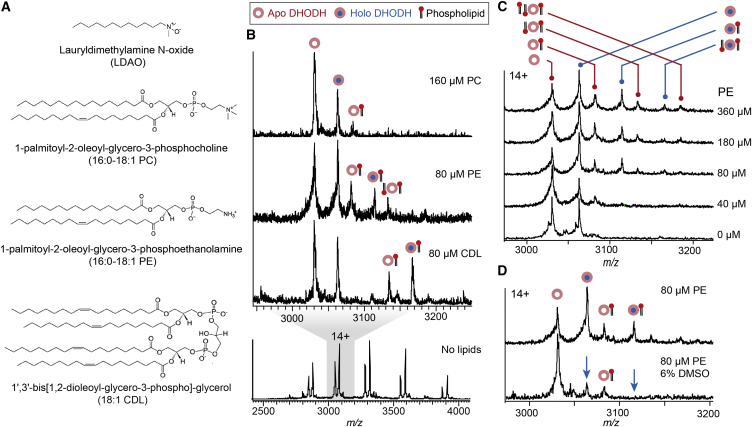
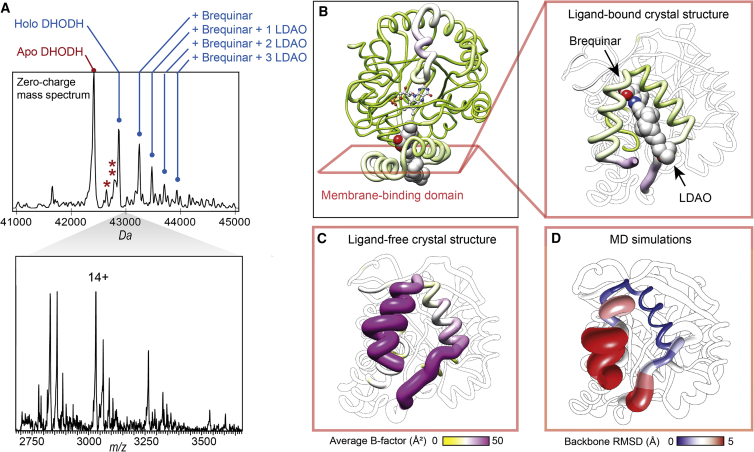
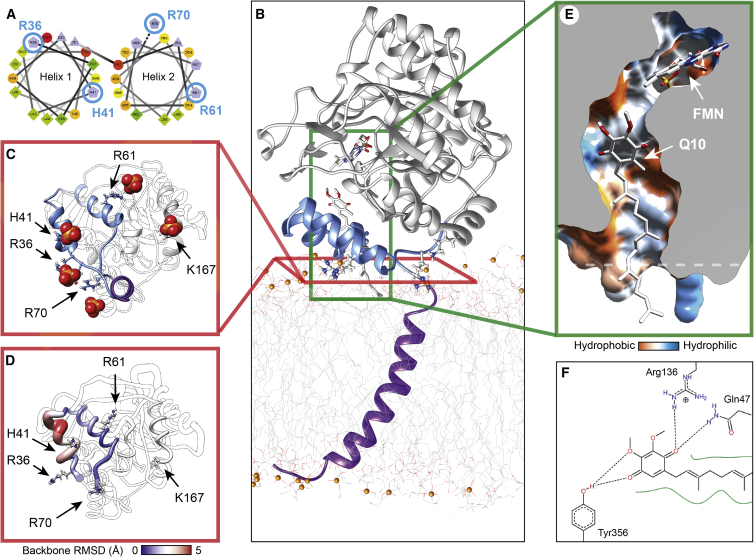
The interactions between proteins and biological membranes are important for drug development, but remain notoriously refractory to structural investigation. We combine non-denaturing mass spectrometry (MS) with molecular dynamics (MD) simulations to unravel the connections among co-factor, lipid, and inhibitor binding in the peripheral membrane protein dihydroorotate dehydrogenase (DHODH), a key anticancer target. Interrogation of intact DHODH complexes by MS reveals that phospholipids bind via their charged head groups at a limited number of sites, while binding of the inhibitor brequinar involves simultaneous association with detergent molecules. MD simulations show that lipids support flexible segments in the membrane-binding domain and position the inhibitor and electron acceptor-binding site away from the membrane surface, similar to the electron acceptor-binding site in respiratory chain complex I. By complementing MS with MD simulations, we demonstrate how a peripheral membrane protein uses lipids to modulate its structure in a similar manner as integral membrane proteins.
Keywords: membrane protein folding; molecular dynamics simulations; non-denaturing mass spectrometry; protein-lipid interactions.
Copyright © 2017 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- Abraham M.J., Murtola T., Schulz R., Páll S., Smith J.C., Hess B., Lindahl E. GROMACS: high performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX. 2015;1–2:19–25.
-
- Angerer H., Nasiri H.R., Niedergesäß V., Kerscher S., Schwalbe H., Brandt U. Tracing the tail of ubiquinone in mitochondrial complex I. Biochim. Biophys. Acta. 2012;1817:1776–1784. - PubMed
-
- Barrera N.P., Di Bartolo N., Booth P.J., Robinson C.V. Micelles protect membrane complexes from solution to vacuum. Science. 2008;321:243–246. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
