

Inhibitory circuit gating of auditory critical-period plasticity
- PMID: 29358666
- PMCID: PMC5978727
- DOI: 10.1038/s41593-017-0064-2
Inhibitory circuit gating of auditory critical-period plasticity
Erratum in
-
Publisher Correction: Inhibitory circuit gating of auditory critical-period plasticity.Nat Neurosci. 2018 Oct;21(10):1495. doi: 10.1038/s41593-018-0161-x. Nat Neurosci. 2018. PMID: 29915196
Abstract
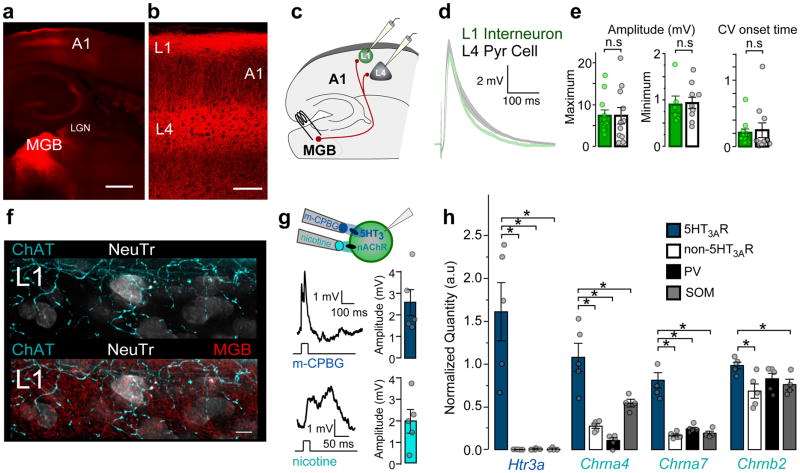
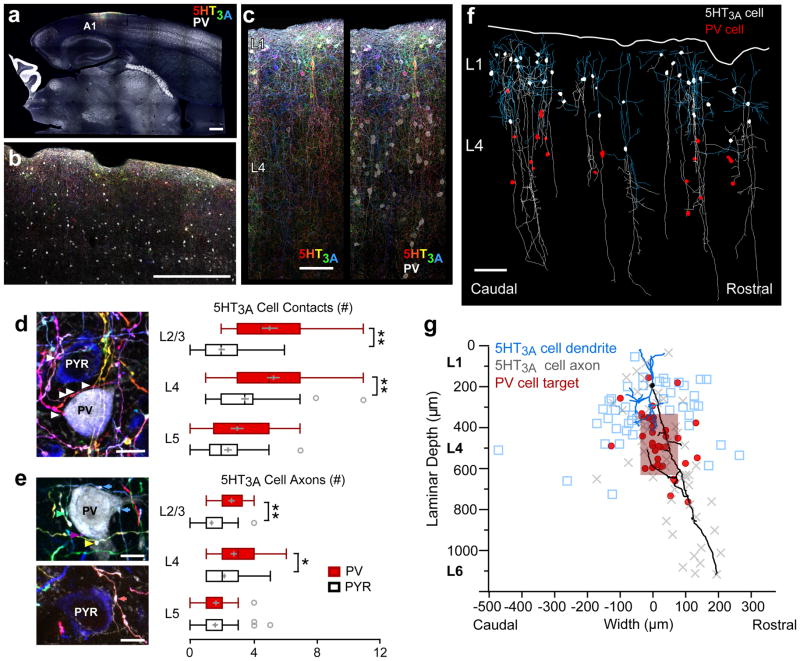
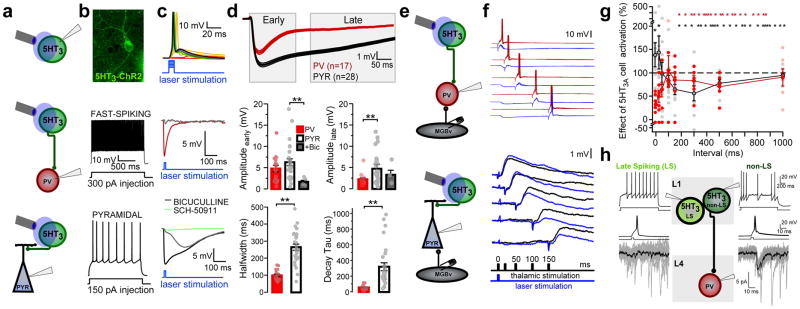
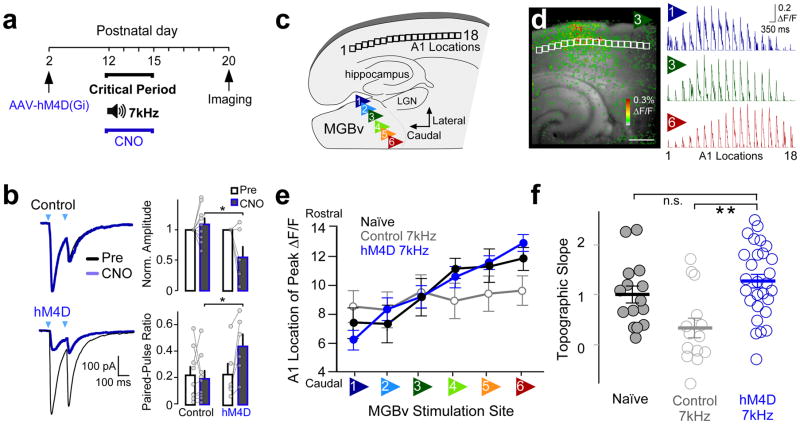
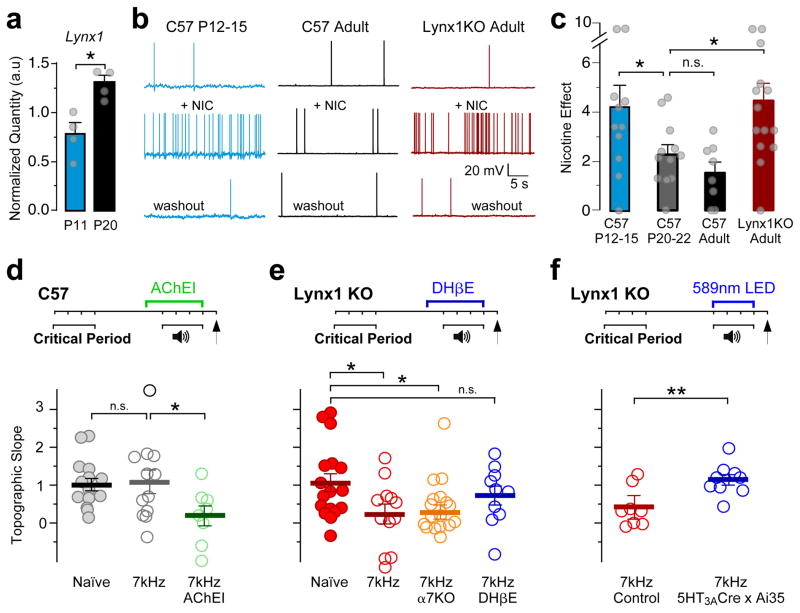
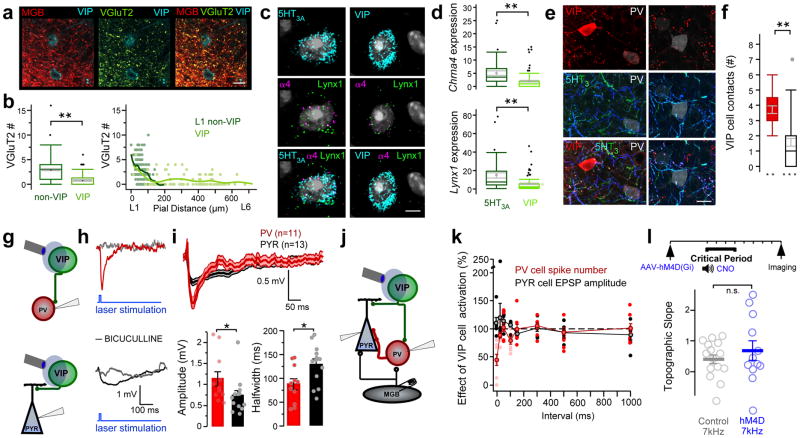
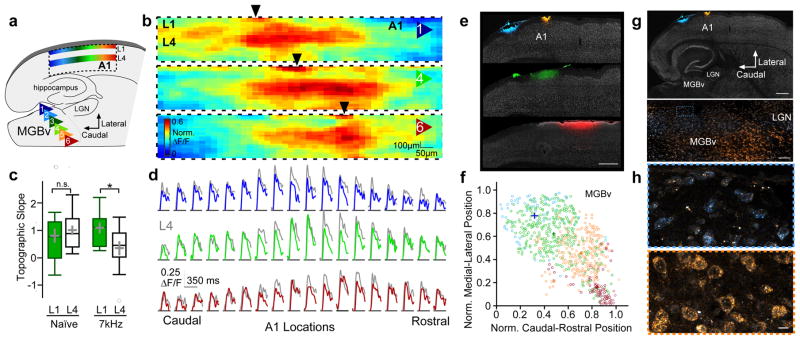
Cortical sensory maps are remodeled during early life to adapt to the surrounding environment. Both sensory and contextual signals are important for induction of this plasticity, but how these signals converge to sculpt developing thalamocortical circuits remains largely unknown. Here we show that layer 1 (L1) of primary auditory cortex (A1) is a key hub where neuromodulatory and topographically organized thalamic inputs meet to tune the cortical layers below. Inhibitory interneurons in L1 send narrowly descending projections to differentially modulate thalamic drive to pyramidal and parvalbumin-expressing (PV) cells in L4, creating brief windows of intracolumnar activation. Silencing of L1 (but not VIP-expressing) cells abolishes map plasticity during the tonotopic critical period. Developmental transitions in nicotinic acetylcholine receptor (nAChR) sensitivity in these cells caused by Lynx1 protein can be overridden to extend critical-period closure. Notably, thalamocortical maps in L1 are themselves stable, and serve as a scaffold for cortical plasticity throughout life.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Werker JF, Hensch TK. Critical Periods in Speech Perception: New Directions. Annual Review Psychol. 2015;66:23.1–23.24. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
