

Fluid network dynamics in the prefrontal cortex during multiple strategy switching
- PMID: 29358717
- PMCID: PMC5778086
- DOI: 10.1038/s41467-017-02764-x
Fluid network dynamics in the prefrontal cortex during multiple strategy switching
Abstract
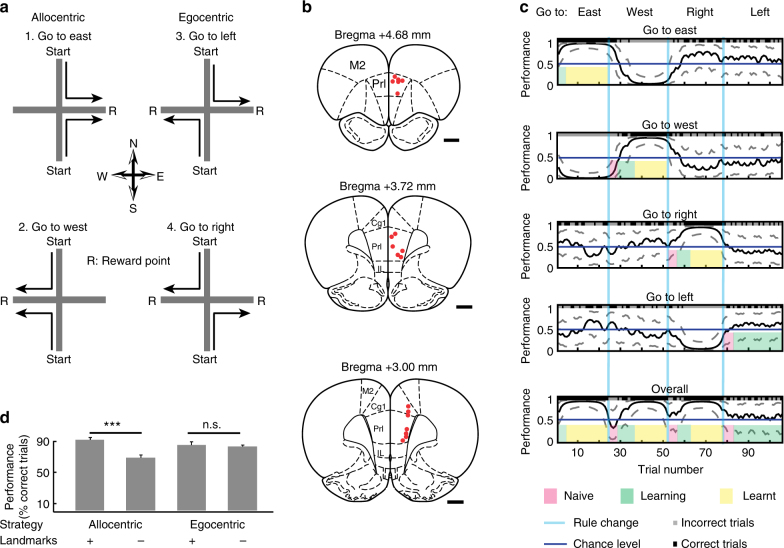
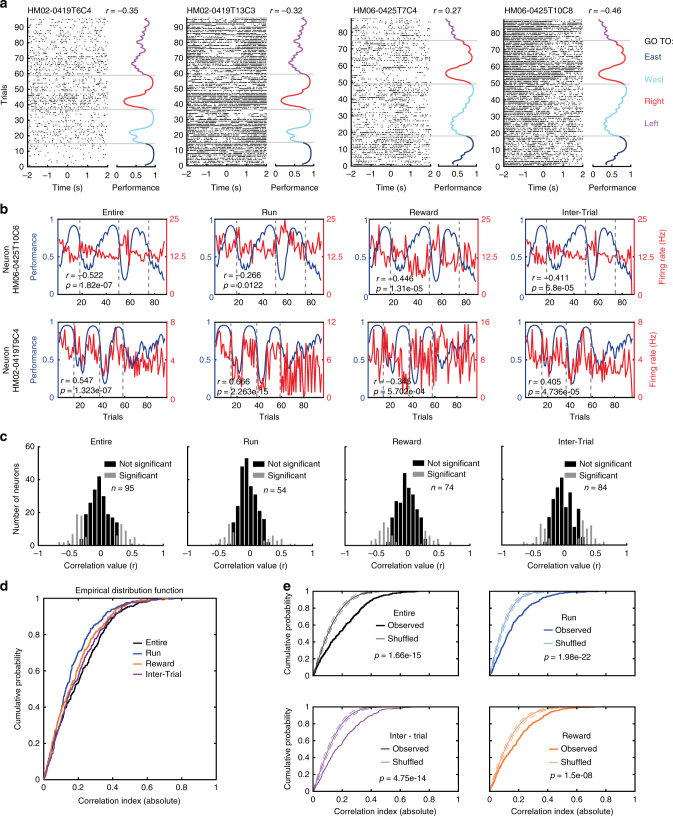
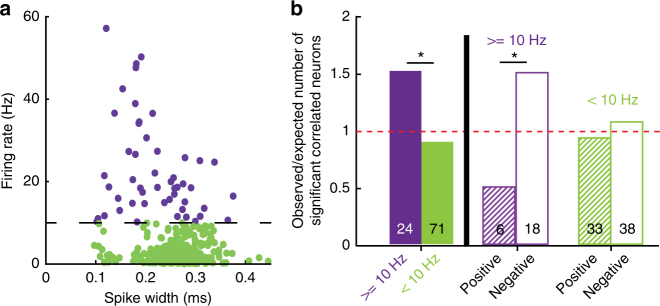
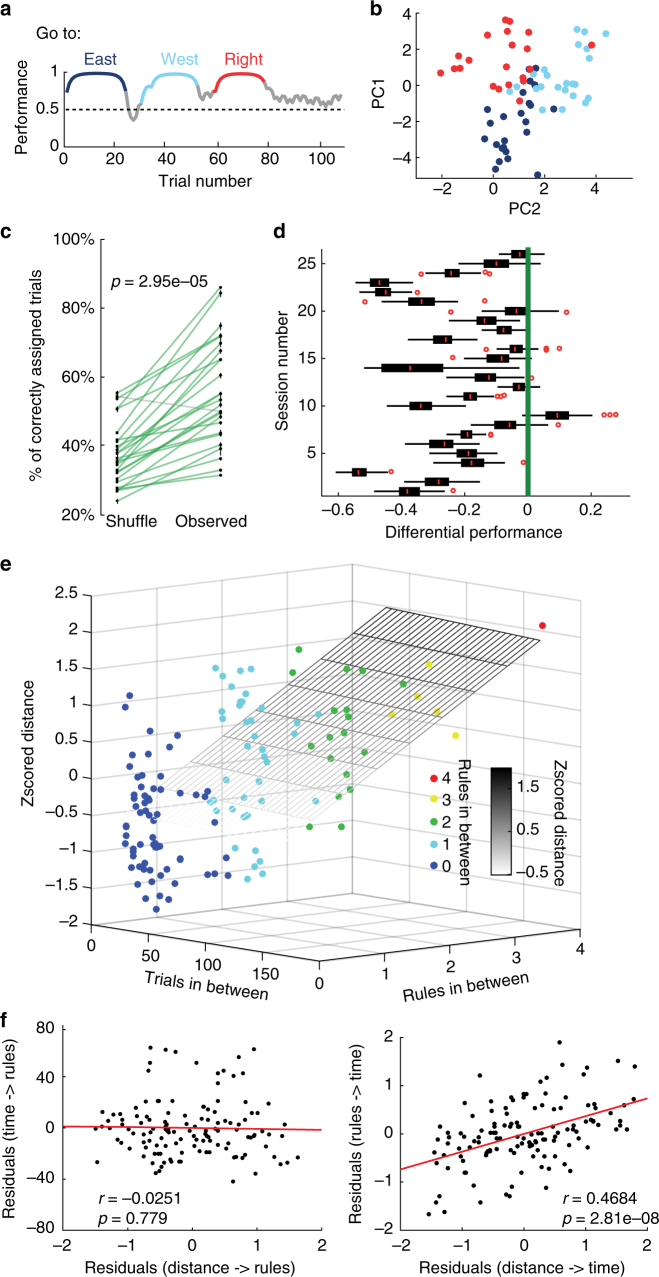
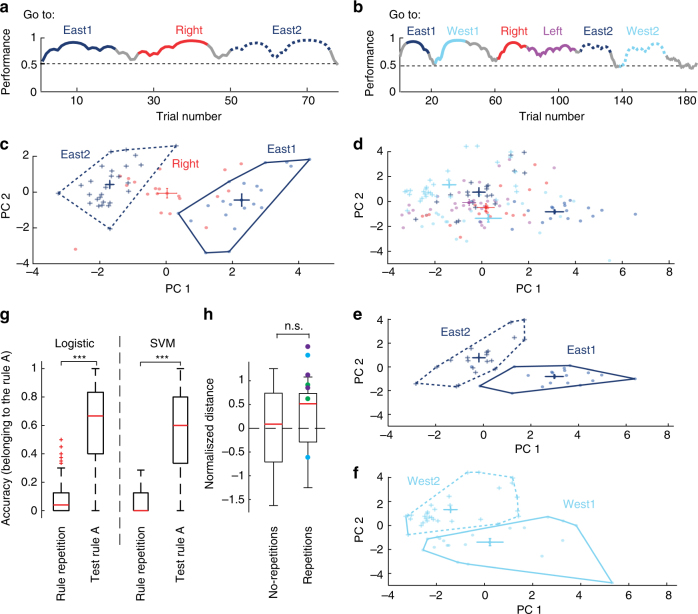
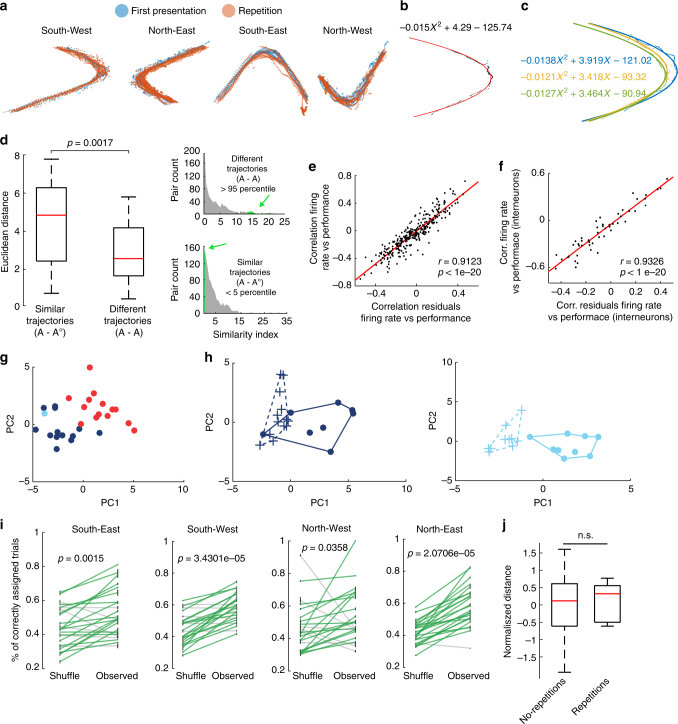
Coordinated shifts of neuronal activity in the prefrontal cortex are associated with strategy adaptations in behavioural tasks, when animals switch from following one rule to another. However, network dynamics related to multiple-rule changes are scarcely known. We show how firing rates of individual neurons in the prelimbic and cingulate cortex correlate with the performance of rats trained to change their navigation multiple times according to allocentric and egocentric strategies. The concerted population activity exhibits a stable firing during the performance of one rule but shifted to another neuronal firing state when a new rule is learnt. Interestingly, when the same rule is presented a second time within the same session, neuronal firing does not revert back to the original neuronal firing state, but a new activity-state is formed. Our data indicate that neuronal firing of prefrontal cortical neurons represents changes in strategy and task-performance rather than specific strategies or rules.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Milner, B. Effects of different brain lesions on card sorting: the role of the frontal lobes. Arch Neurol. 9(1), 90–100 (1963).
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
