

Feeding Immunity: Physiological and Behavioral Responses to Infection and Resource Limitation
- PMID: 29358937
- PMCID: PMC5766659
- DOI: 10.3389/fimmu.2017.01914
Feeding Immunity: Physiological and Behavioral Responses to Infection and Resource Limitation
Abstract
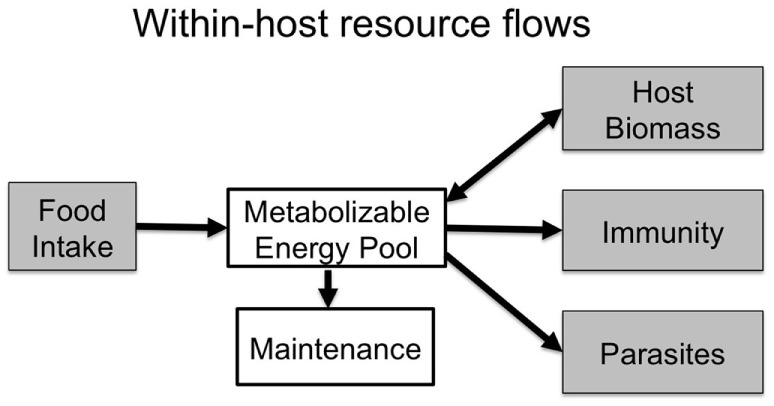
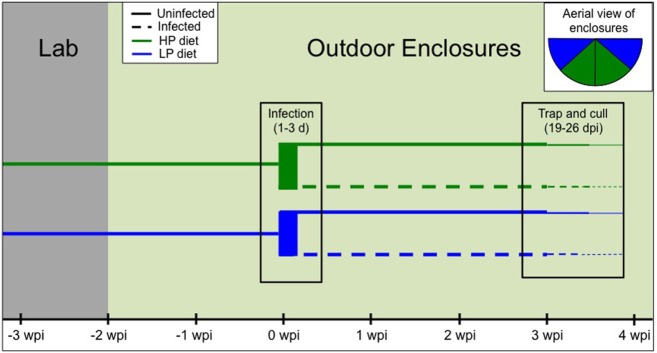
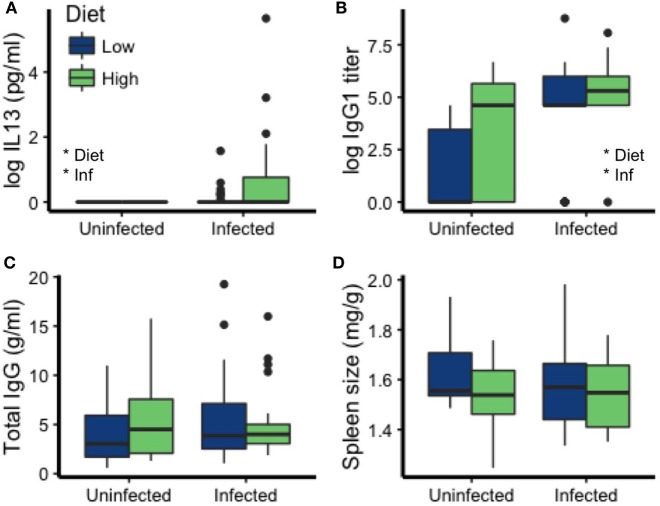
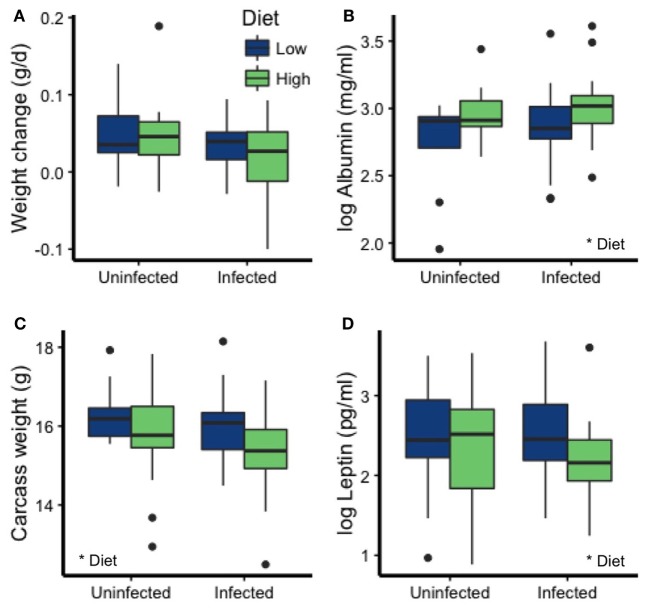
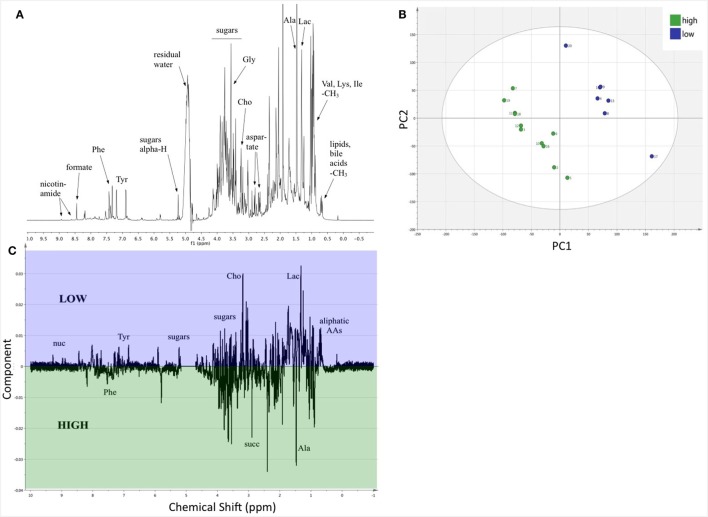
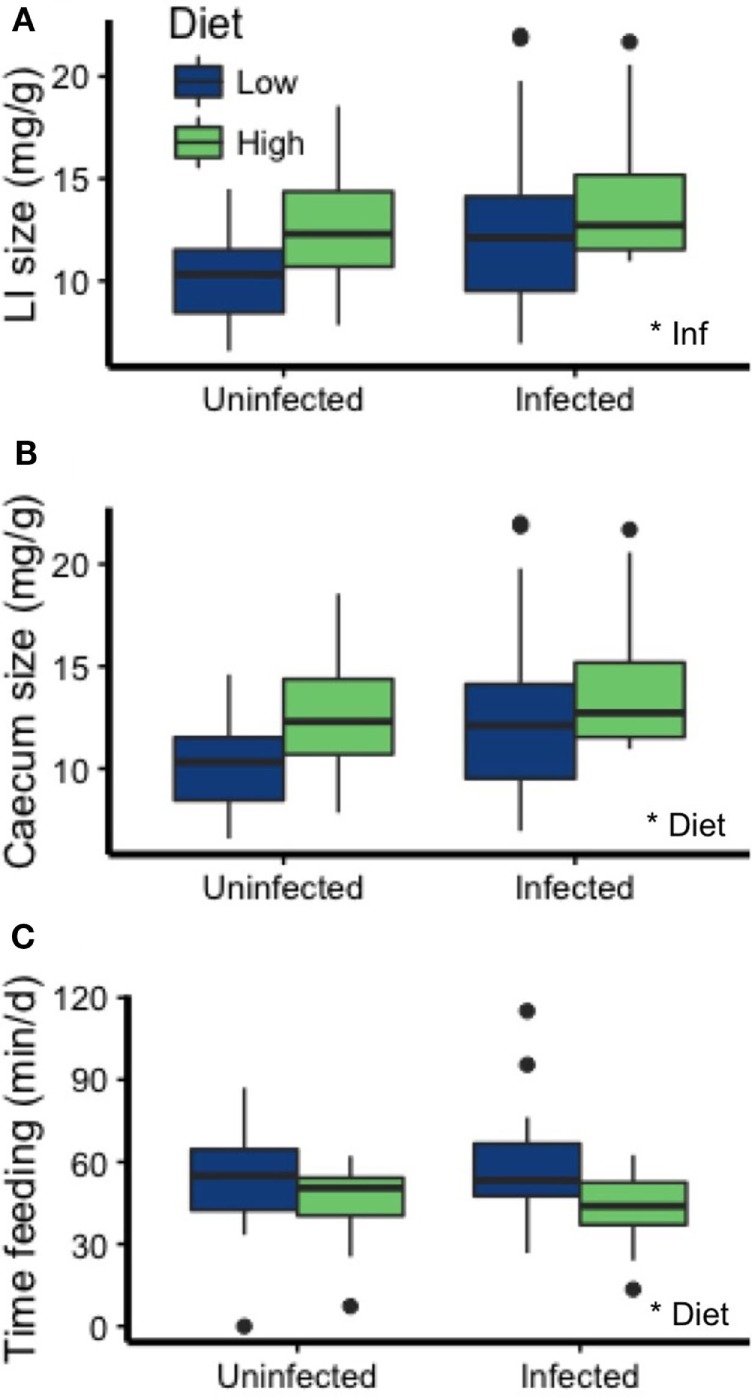
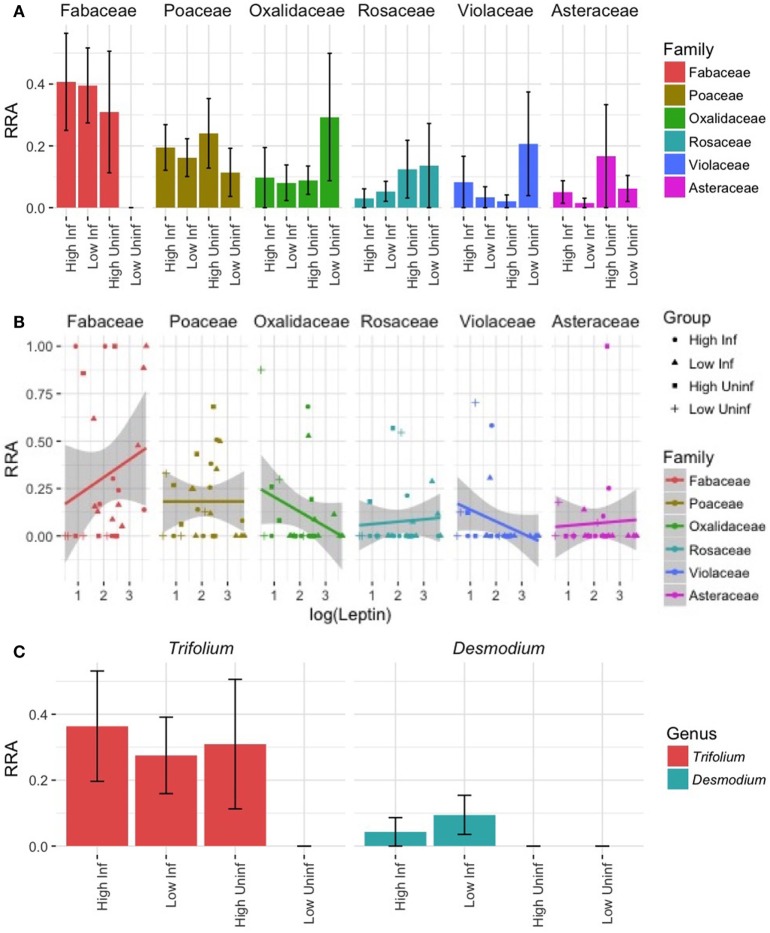
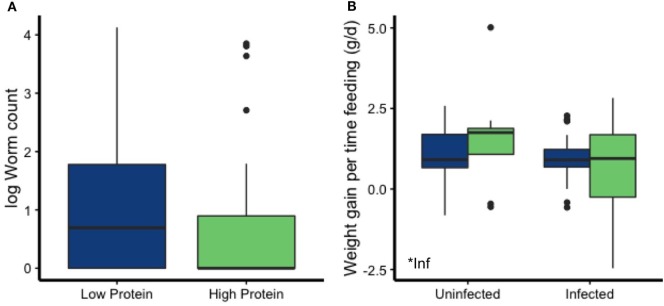
Resources are a core currency of species interactions and ecology in general (e.g., think of food webs or competition). Within parasite-infected hosts, resources are divided among the competing demands of host immunity and growth as well as parasite reproduction and growth. Effects of resources on immune responses are increasingly understood at the cellular level (e.g., metabolic predictors of effector function), but there has been limited consideration of how these effects scale up to affect individual energetic regimes (e.g., allocation trade-offs), susceptibility to infection, and feeding behavior (e.g., responses to local resource quality and quantity). We experimentally rewilded laboratory mice (strain C57BL/6) in semi-natural enclosures to investigate the effects of dietary protein and gastrointestinal nematode (Trichuris muris) infection on individual-level immunity, activity, and behavior. The scale and realism of this field experiment, as well as the multiple physiological assays developed for laboratory mice, enabled us to detect costs, trade-offs, and potential compensatory mechanisms that mice employ to battle infection under different resource conditions. We found that mice on a low-protein diet spent more time feeding, which led to higher body fat stores (i.e., concentration of a satiety hormone, leptin) and altered metabolite profiles, but which did not fully compensate for the effects of poor nutrition on albumin or immune defenses. Specifically, immune defenses measured as interleukin 13 (IL13) (a primary cytokine coordinating defense against T. muris) and as T. muris-specific IgG1 titers were lower in mice on the low-protein diet. However, these reduced defenses did not result in higher worm counts in mice with poorer diets. The lab mice, living outside for the first time in thousands of generations, also consumed at least 26 wild plant species occurring in the enclosures, and DNA metabarcoding revealed that the consumption of different wild foods may be associated with differences in leptin concentrations. When individual foraging behavior was accounted for, worm infection significantly reduced rates of host weight gain. Housing laboratory mice in outdoor enclosures provided new insights into the resource costs of immune defense to helminth infection and how hosts modify their behavior to compensate for those costs.
Keywords: DNA metabarcoding; Trichuris muris; compensatory feeding; nuclear magnetic resonance spectroscopy metabolite profiling; resource–immune trade-offs; rewilding mice.
Figures
References
-
- Fittkau EJ, Klinge H. On biomass and trophic structure of the Central Amazonian rain forest ecosystem. Biotropica (1973) 5:2–14. 10.2307/2989676 - DOI
-
- McNaughton SJ. Ecology of a grazing ecosystem: the Serengeti. Ecol Monogr (1985) 55:260–94. 10.2307/1942578 - DOI
-
- Lindeman RL. The trophic-dynamic aspect of ecology. Ecology (1942) 23:399–417. 10.2307/1930126 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous