

Structure of the human activated spliceosome in three conformational states
- PMID: 29360106
- PMCID: PMC5835773
- DOI: 10.1038/cr.2018.14
Structure of the human activated spliceosome in three conformational states
Abstract
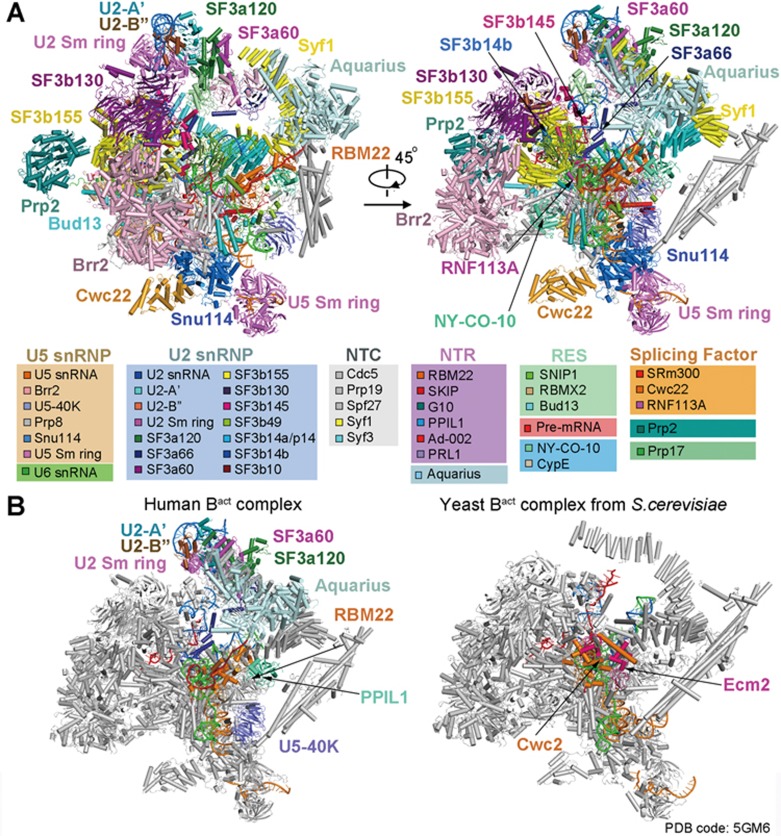
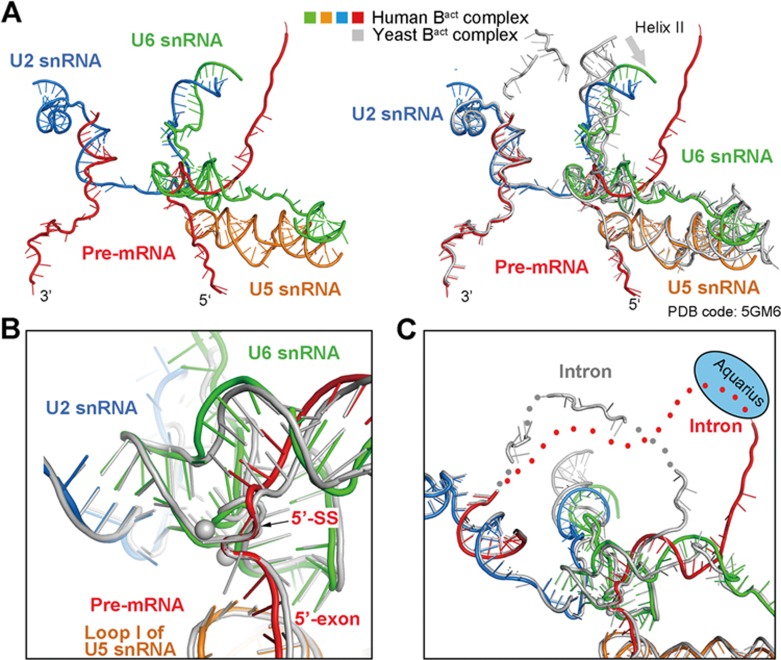
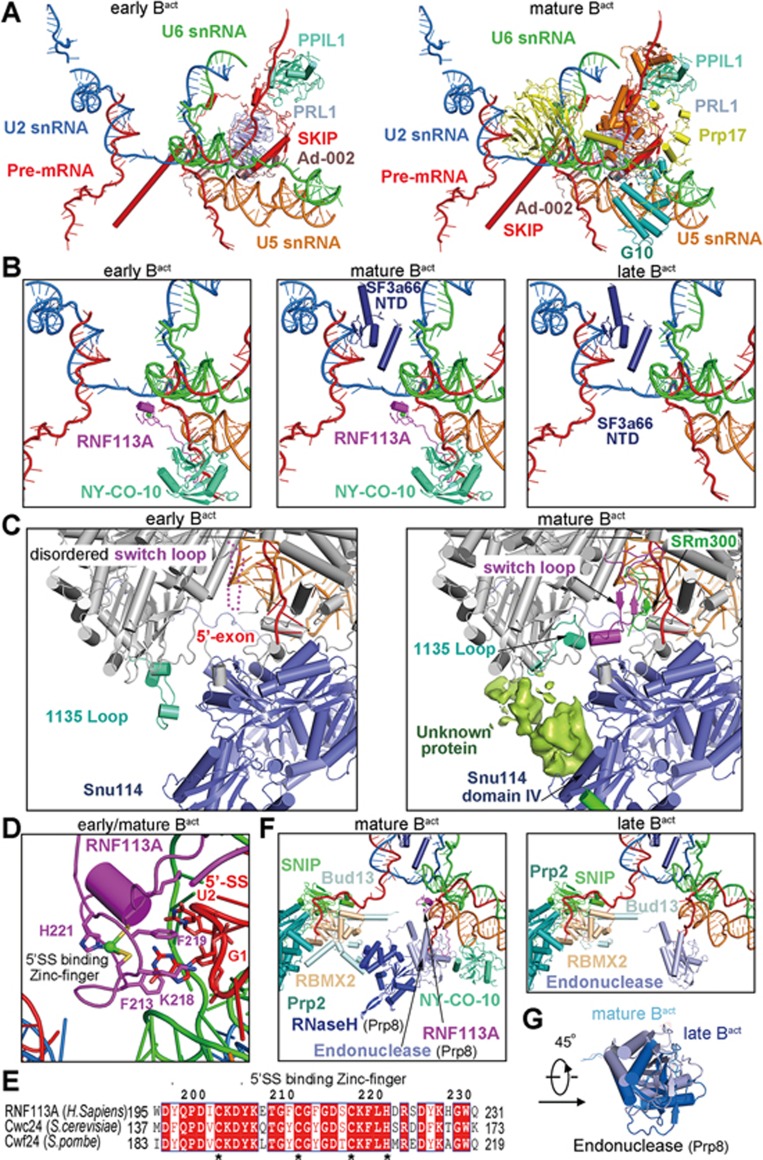
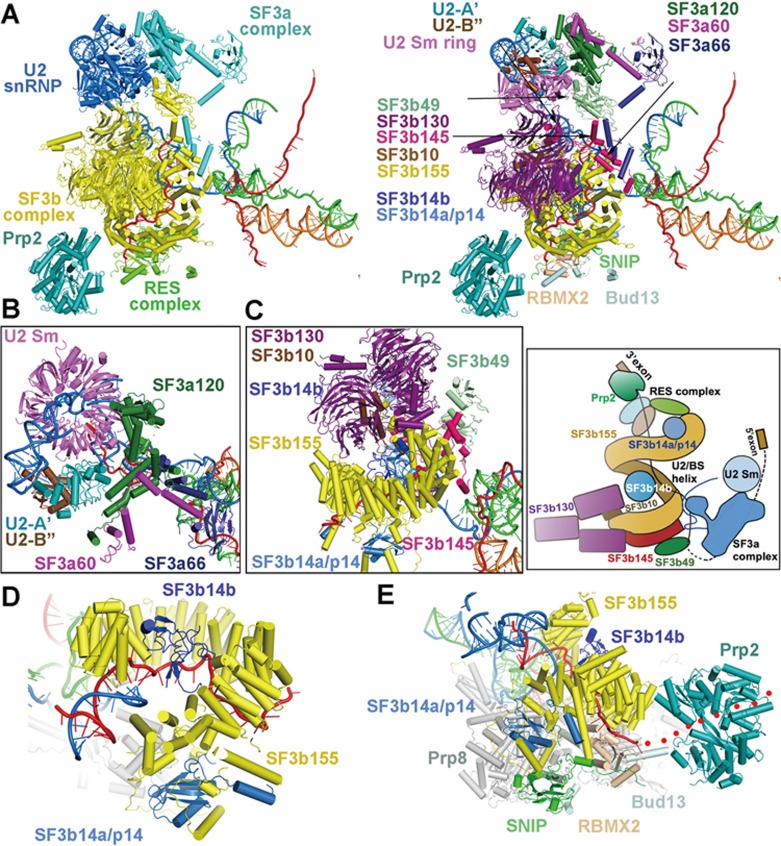
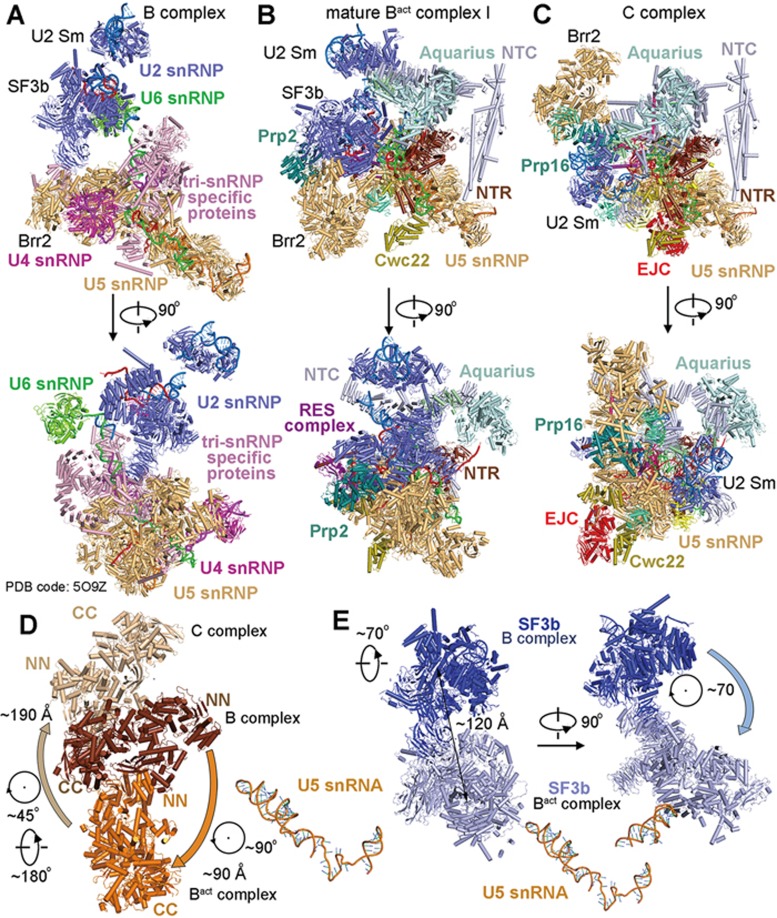
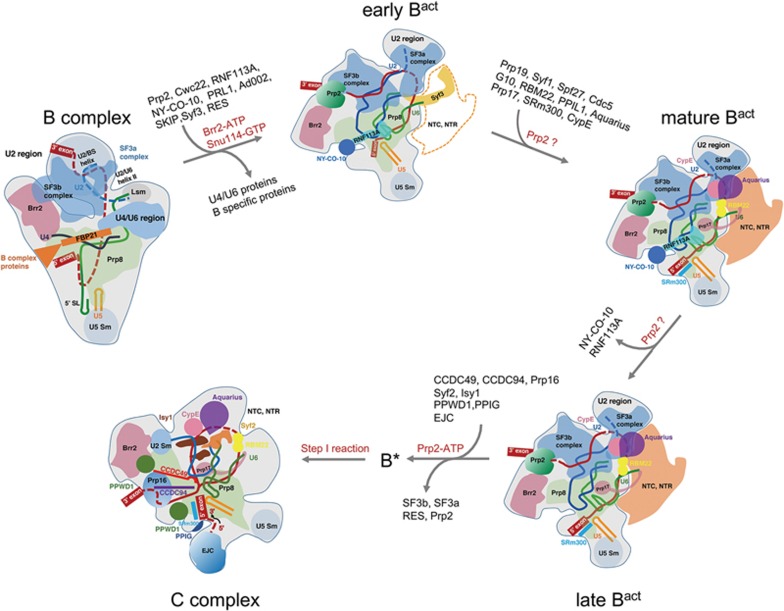
During each cycle of pre-mRNA splicing, the pre-catalytic spliceosome (B complex) is converted into the activated spliceosome (Bact complex), which has a well-formed active site but cannot proceed to the branching reaction. Here, we present the cryo-EM structure of the human Bact complex in three distinct conformational states. The EM map allows atomic modeling of nearly all protein components of the U2 small nuclear ribonucleoprotein (snRNP), including three of the SF3a complex and seven of the SF3b complex. The structure of the human Bact complex contains 52 proteins, U2, U5, and U6 small nuclear RNA (snRNA), and a pre-mRNA. Three distinct conformations have been captured, representing the early, mature, and late states of the human Bact complex. These complexes differ in the orientation of the Switch loop of Prp8, the splicing factors RNF113A and NY-CO-10, and most components of the NineTeen complex (NTC) and the NTC-related complex. Analysis of these three complexes and comparison with the B and C complexes reveal an ordered flux of components in the B-to-Bact and the Bact-to-B* transitions, which ultimately prime the active site for the branching reaction.
Figures
References
-
- Wahl MC, Will CL, Luhrmann R. The spliceosome: design principles of a dynamic RNP machine. Cell 2009; 136:701–718. - PubMed
-
- Chan SP, Kao DI, Tsai WY, Cheng SC. The Prp19p-associated complex in spliceosome activation. Science 2003; 302:279–282. - PubMed
-
- Fabrizio P, Dannenberg J, Dube P, et al. The evolutionarily conserved core design of the catalytic activation step of the yeast spliceosome. Mol Cell 2009; 36:593–608. - PubMed
-
- Yan C, Wan R, Bai R, Huang G, Shi Y. Structure of a yeast activated spliceosome at 3.5 A resolution. Science 2016; 353:904–911. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
