

A sigma factor toolbox for orthogonal gene expression in Escherichia coli
- PMID: 29361130
- PMCID: PMC5829568
- DOI: 10.1093/nar/gky010
A sigma factor toolbox for orthogonal gene expression in Escherichia coli
Abstract
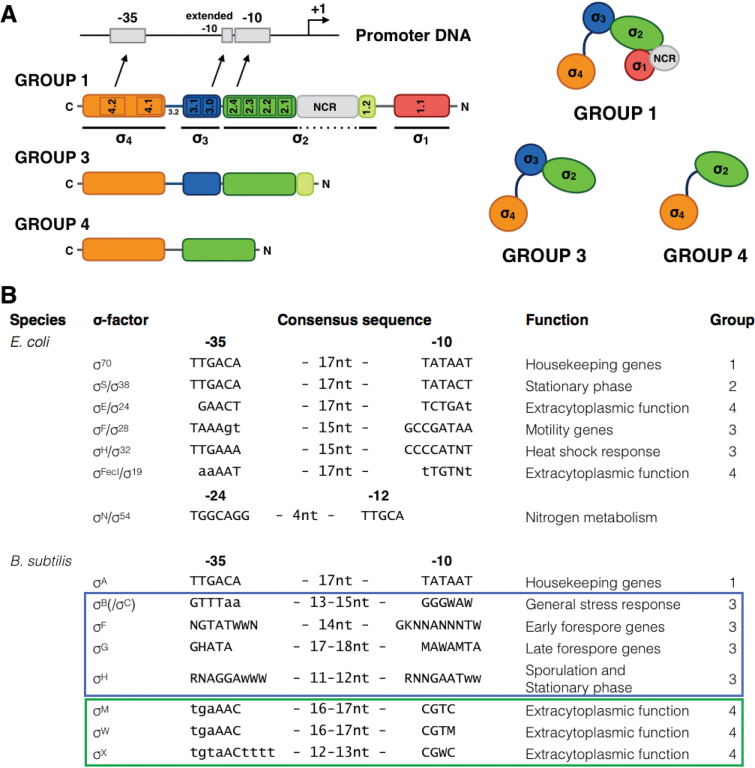
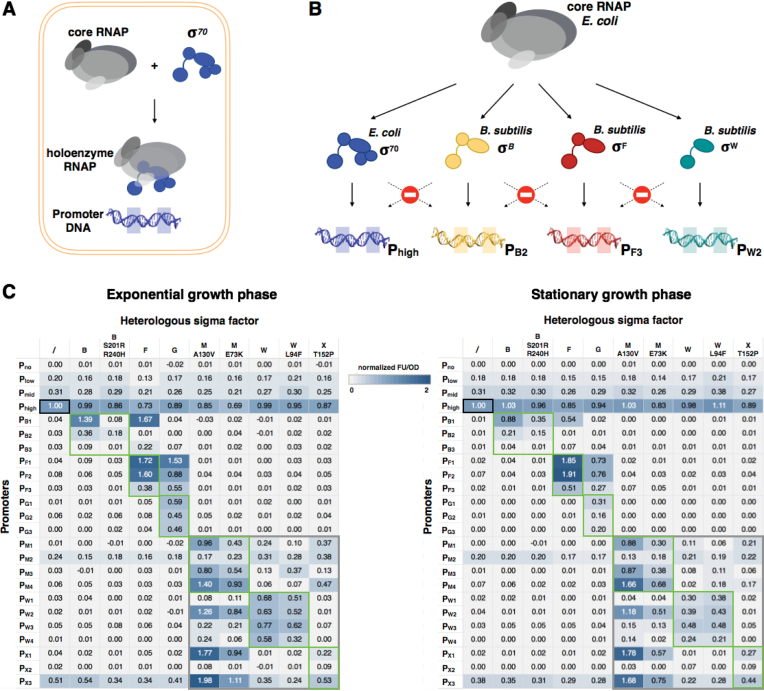
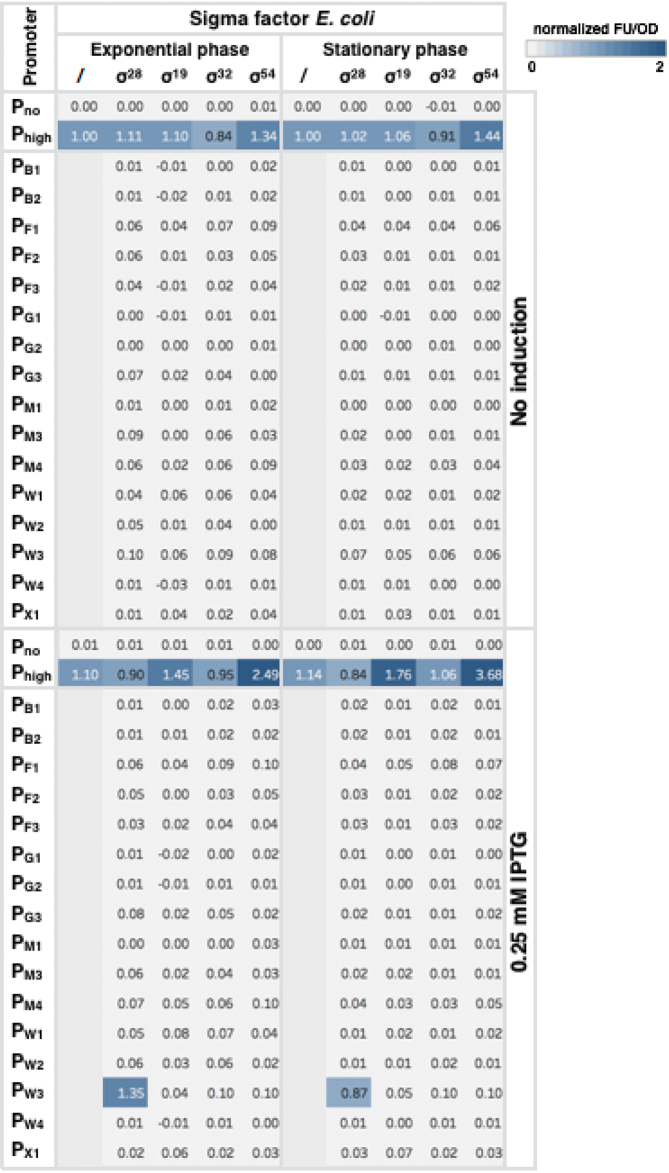
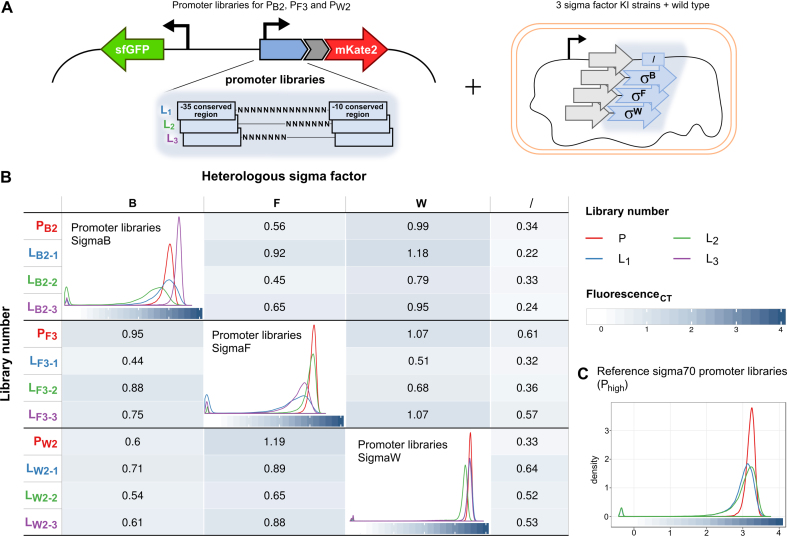
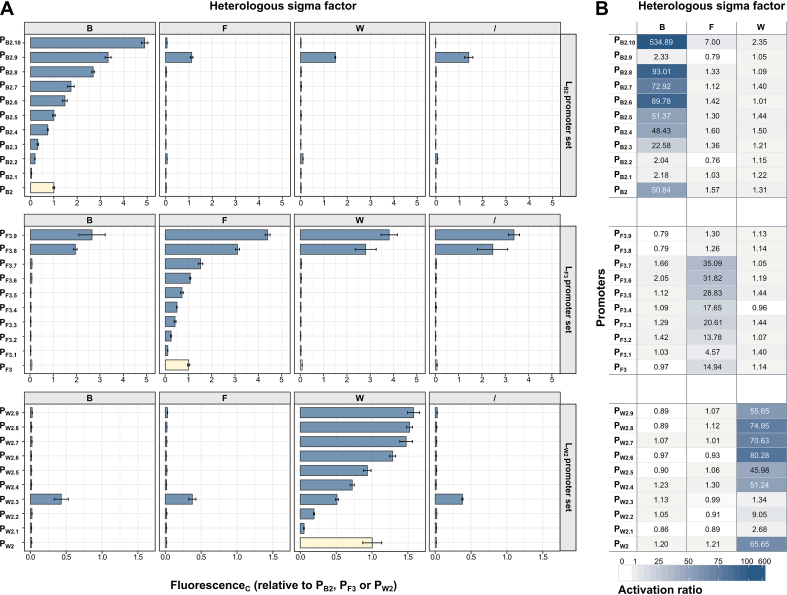
Synthetic genetic sensors and circuits enable programmable control over timing and conditions of gene expression and, as a result, are increasingly incorporated into the control of complex and multi-gene pathways. Size and complexity of genetic circuits are growing, but stay limited by a shortage of regulatory parts that can be used without interference. Therefore, orthogonal expression and regulation systems are needed to minimize undesired crosstalk and allow for dynamic control of separate modules. This work presents a set of orthogonal expression systems for use in Escherichia coli based on heterologous sigma factors from Bacillus subtilis that recognize specific promoter sequences. Up to four of the analyzed sigma factors can be combined to function orthogonally between each other and toward the host. Additionally, the toolbox is expanded by creating promoter libraries for three sigma factors without loss of their orthogonal nature. As this set covers a wide range of transcription initiation frequencies, it enables tuning of multiple outputs of the circuit in response to different sensory signals in an orthogonal manner. This sigma factor toolbox constitutes an interesting expansion of the synthetic biology toolbox and may contribute to the assembly of more complex synthetic genetic systems in the future.
Figures
References
-
- Wargacki A.J., Leonard E., Win M.N., Regitsky D.D., Santos C.N.S., Kim P.B., Cooper S.R., Raisner R.M., Herman A., Sivitz A.B. et al. . An engineered microbial platform for direct biofuel production from brown macroalgae. Science. 2012; 335:308–313. - PubMed
-
- Cheon S., Kim H.M., Gustavsson M., Lee S.Y.. Recent trends in metabolic engineering of microorganisms for the production of advanced biofuels. Curr. Opin. Chem. Biol. 2016; 35:10–21. - PubMed
-
- Pandey R.P., Parajuli P., Koffas M.A.G., Sohng J.K.. Microbial production of natural and non-natural flavonoids: Pathway engineering, directed evolution and systems/synthetic biology. Biotechnol. Adv. 2016; 34:634–662. - PubMed
-
- Paddon C.J., Keasling J.D.. Semi-synthetic artemisinin: a model for the use of synthetic biology in pharmaceutical development. Nat. Rev. Microbiol. 2014; 12:355–367. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
