

Septins are involved at the early stages of macroautophagy in S. cerevisiae
- PMID: 29361537
- PMCID: PMC5868950
- DOI: 10.1242/jcs.209098
Septins are involved at the early stages of macroautophagy in S. cerevisiae
Abstract
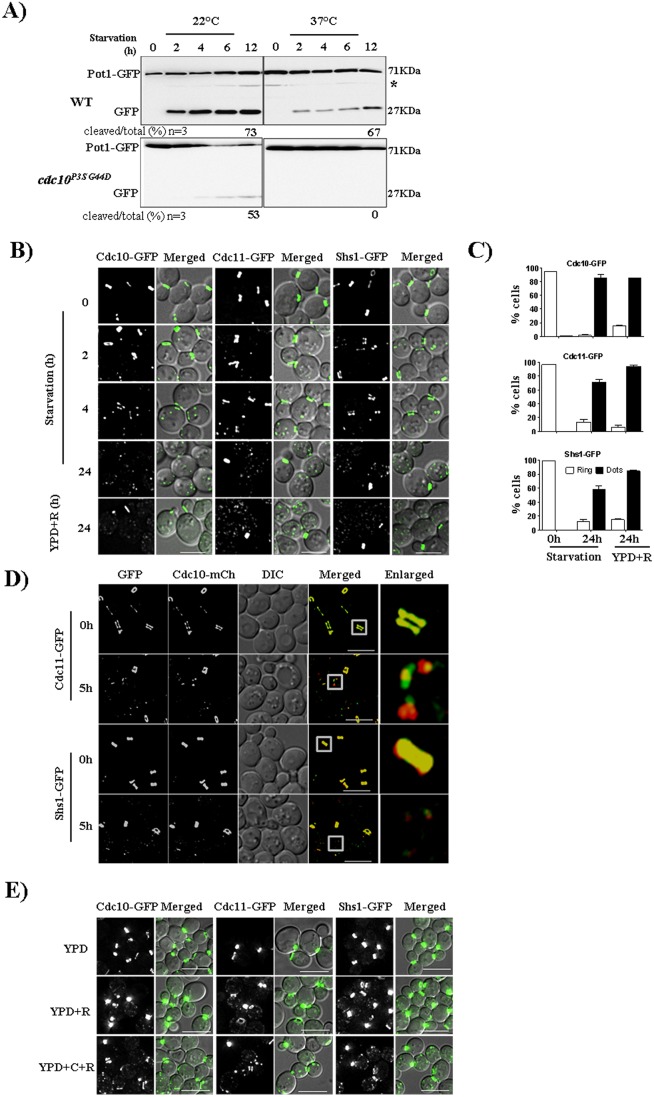
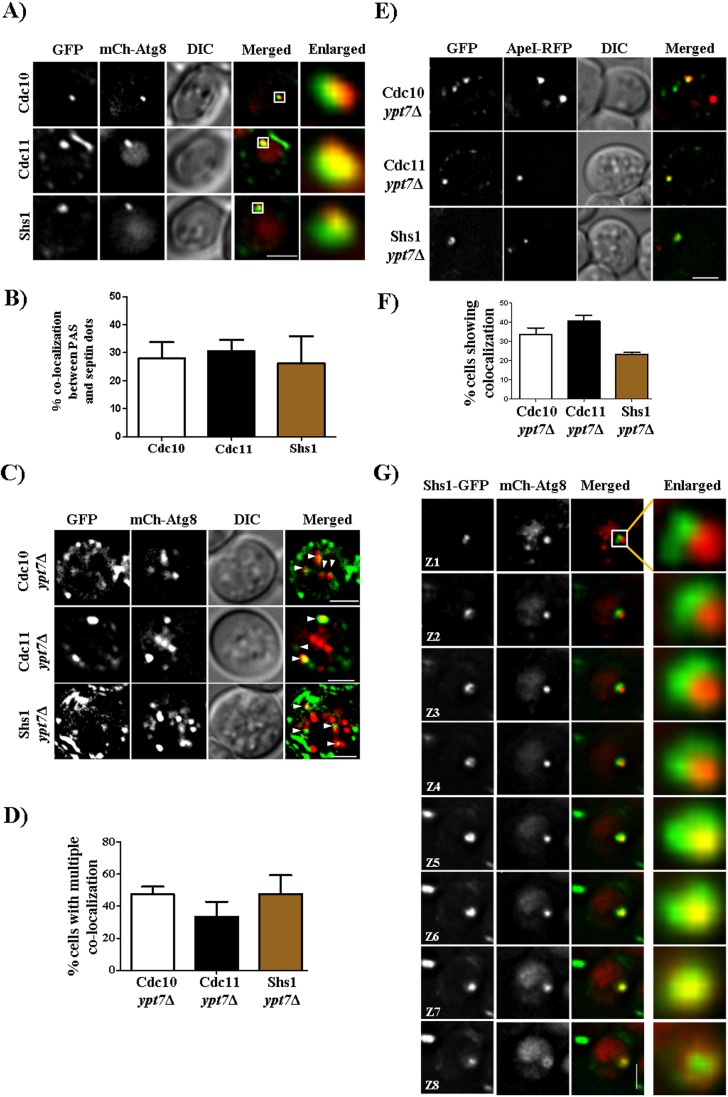
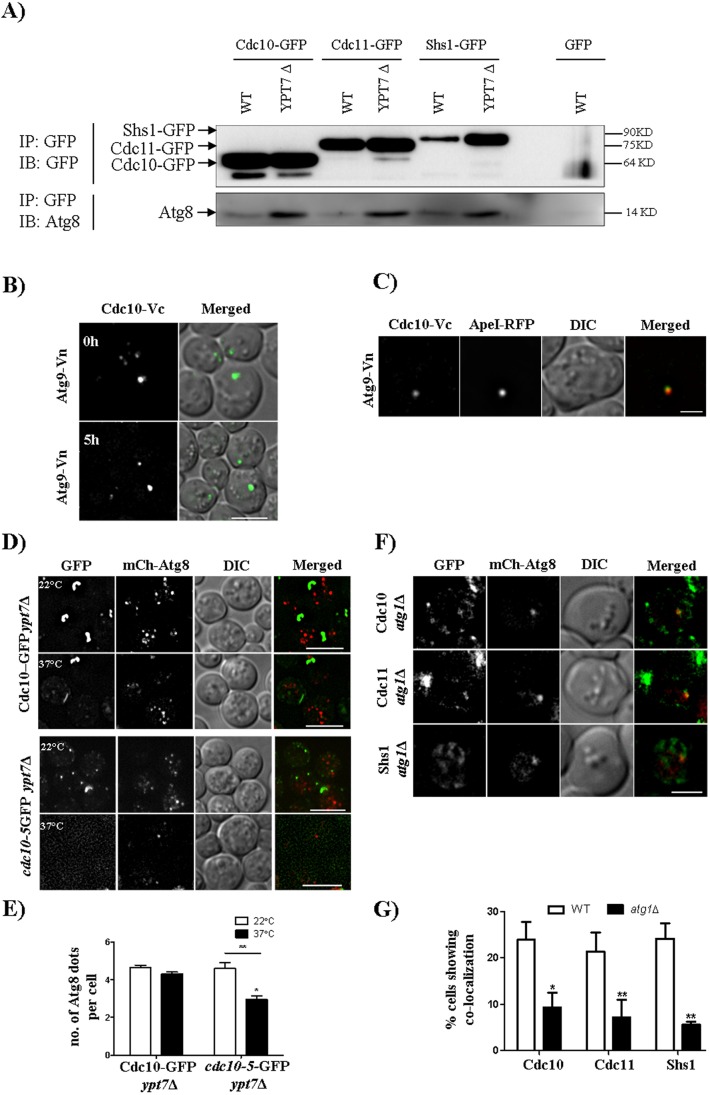
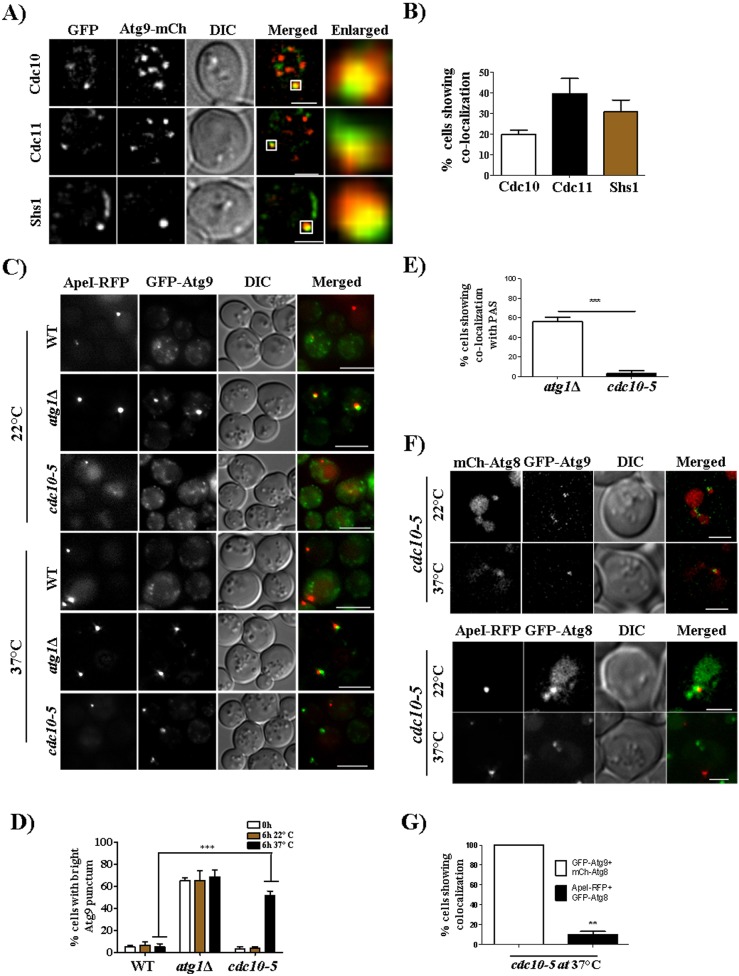
Autophagy is a conserved cellular degradation pathway wherein double-membrane vesicles called autophagosomes capture long-lived proteins, and damaged or superfluous organelles, and deliver them to the lysosome for degradation. Septins are conserved GTP-binding proteins involved in many cellular processes, including phagocytosis and the autophagy of intracellular bacteria, but no role in general autophagy was known. In budding yeast, septins polymerize into ring-shaped arrays of filaments required for cytokinesis. In an unbiased genetic screen and in subsequent targeted analysis, we found autophagy defects in septin mutants. Upon autophagy induction, pre-assembled septin complexes relocalized to the pre-autophagosomal structure (PAS) where they formed non-canonical septin rings at PAS. Septins also colocalized with autophagosomes, where they physically interacted with the autophagy proteins Atg8 and Atg9. When autophagosome degradation was blocked in septin-mutant cells, fewer autophagic structures accumulated, and an autophagy mutant defective in early stages of autophagosome biogenesis (atg1Δ), displayed decreased septin localization to the PAS. Our findings support a role for septins in the early stages of budding yeast autophagy, during autophagosome formation.This article has an associated First Person interview with the first author of the paper.
Keywords: Atg9 trafficking; Autophagosome biogenesis; Autophagy; Noncanonical ring; PAS; Pre-autophagosomal structure; Septin.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
