

Critical neutralizing fragment of Zika virus EDIII elicits cross-neutralization and protection against divergent Zika viruses
- PMID: 29362446
- PMCID: PMC5837162
- DOI: 10.1038/s41426-017-0007-8
Critical neutralizing fragment of Zika virus EDIII elicits cross-neutralization and protection against divergent Zika viruses
Abstract
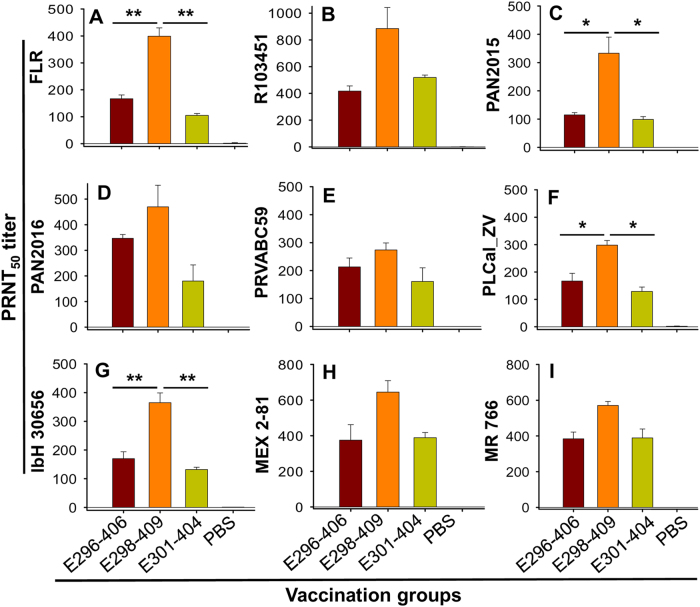
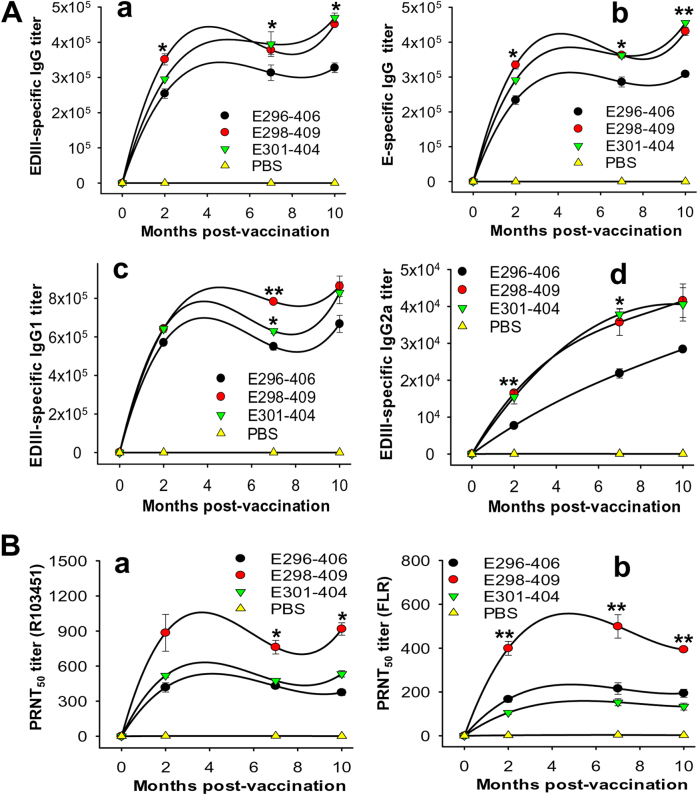
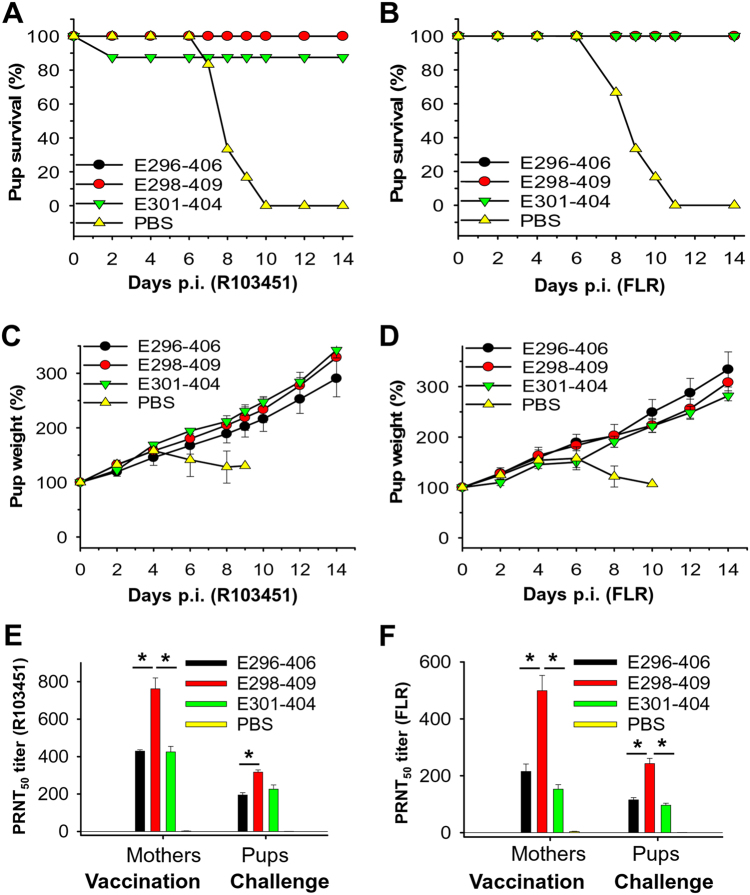
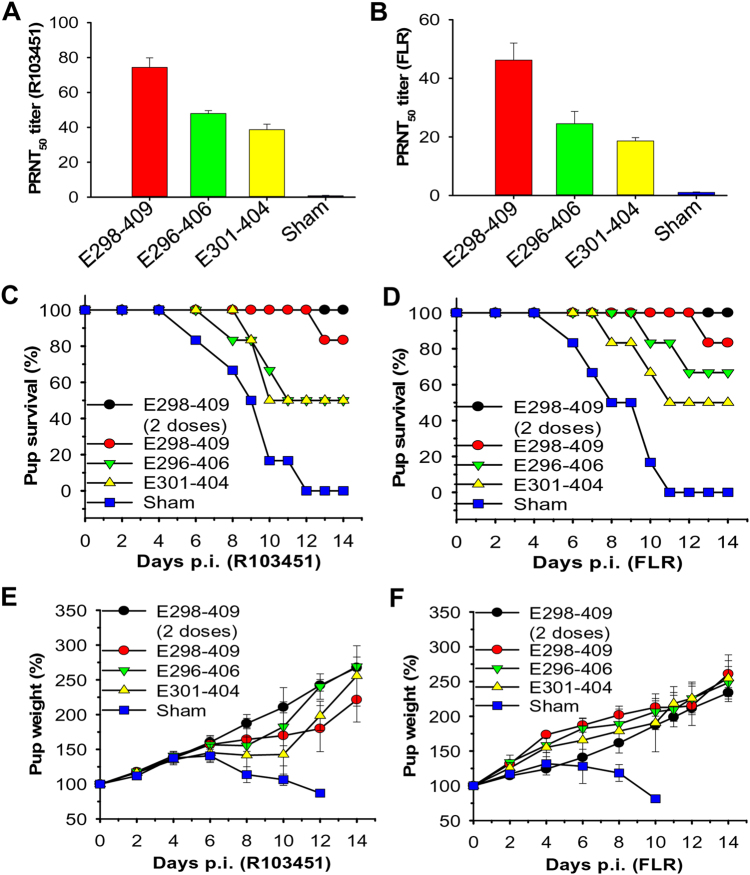
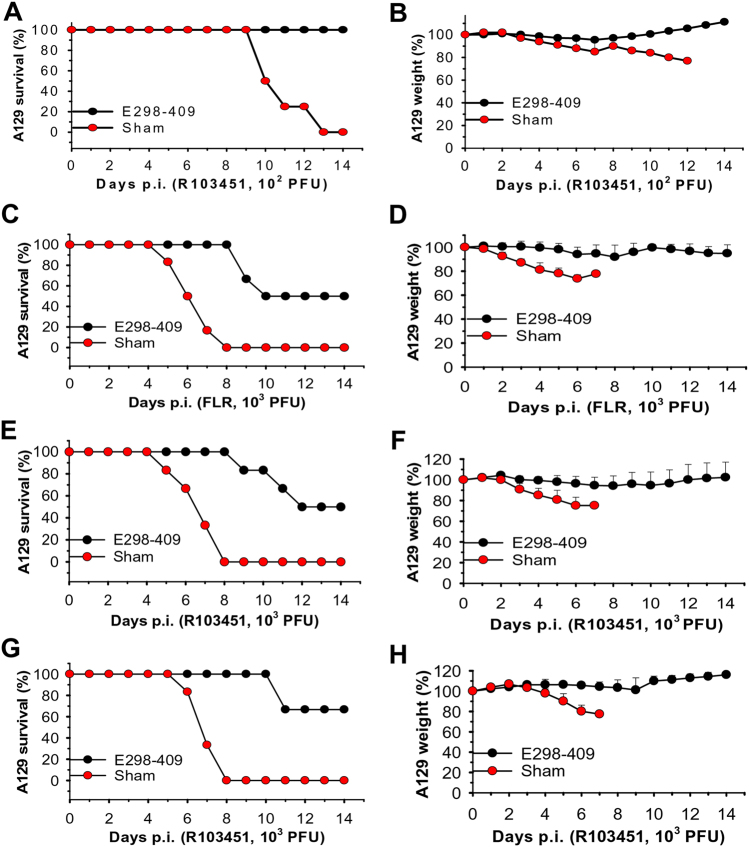
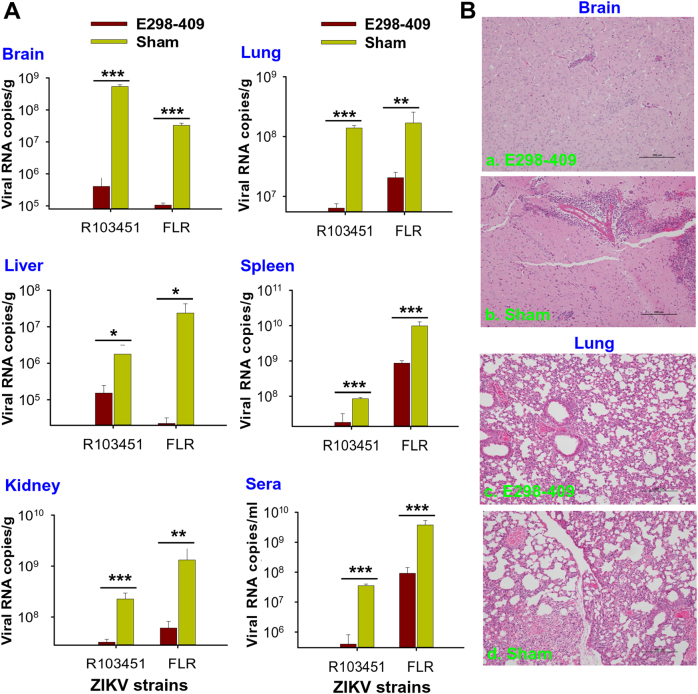
Zika virus (ZIKV) infection remains a serious health threat due to its close association with congenital Zika syndrome (CZS), which includes microcephaly and other severe birth defects. As no vaccines are available for human use, continuous effort is needed to develop effective and safe vaccines to prevent ZIKV infection. In this study, we constructed three recombinant proteins comprising, respectively, residues 296-406 (E296-406), 298-409 (E298-409), and 301-404 (E301-404) of ZIKV envelope (E) protein domain III (EDIII) fused with a C-terminal Fc of human IgG. Our results demonstrated that E298-409 induced the highest titer of neutralizing antibodies against infection with nine ZIKV strains isolated from different hosts, countries, and time periods, and it maintained long-term anti-ZIKV immunogenicity to induce neutralizing antibodies. Pups born to mice immunized with E298-409 were fully protected against lethal challenge with two epidemic human ZIKV strains, 2015/Honduras (R103451) and 2015/Colombia (FLR). Passive transfer of anti-E298-409 mouse sera protected pups born to naive mice, as well as type I interferon receptor-deficient adult A129 mice, from lethal challenge with human ZIKV strains R103451 and FLR, and this protection was positively correlated with neutralizing antibodies. These data suggest that the critical neutralizing fragment (i.e., a fragment that can induce highly potent neutralizing antibodies against divergent ZIKV strains) of ZIKV EDIII is a good candidate for development as an effective and safe ZIKV subunit vaccine to protect pregnant mothers and their fetuses against ZIKV infection. The E298-409-specific antibodies can be used for passive immunization to prevent ZIKV infection in newborns or immunocompromised adults.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical