

Mitochondria are transported along microtubules in membrane nanotubes to rescue distressed cardiomyocytes from apoptosis
- PMID: 29362447
- PMCID: PMC5833423
- DOI: 10.1038/s41419-017-0145-x
Mitochondria are transported along microtubules in membrane nanotubes to rescue distressed cardiomyocytes from apoptosis
Abstract
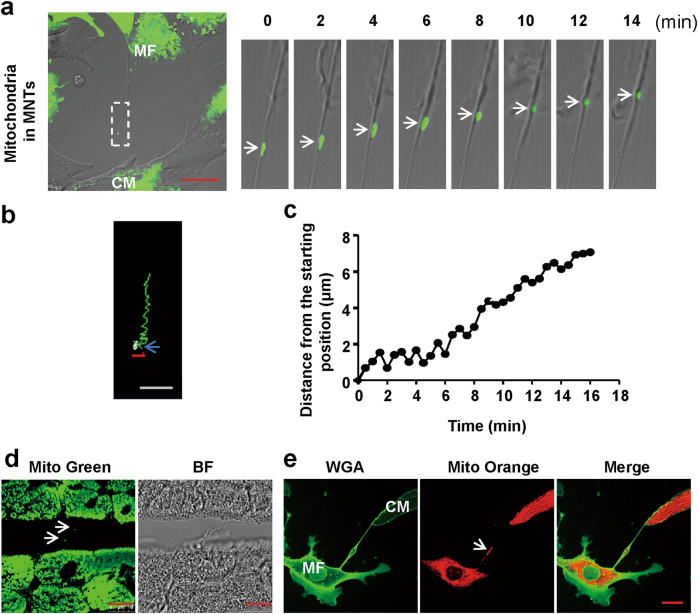
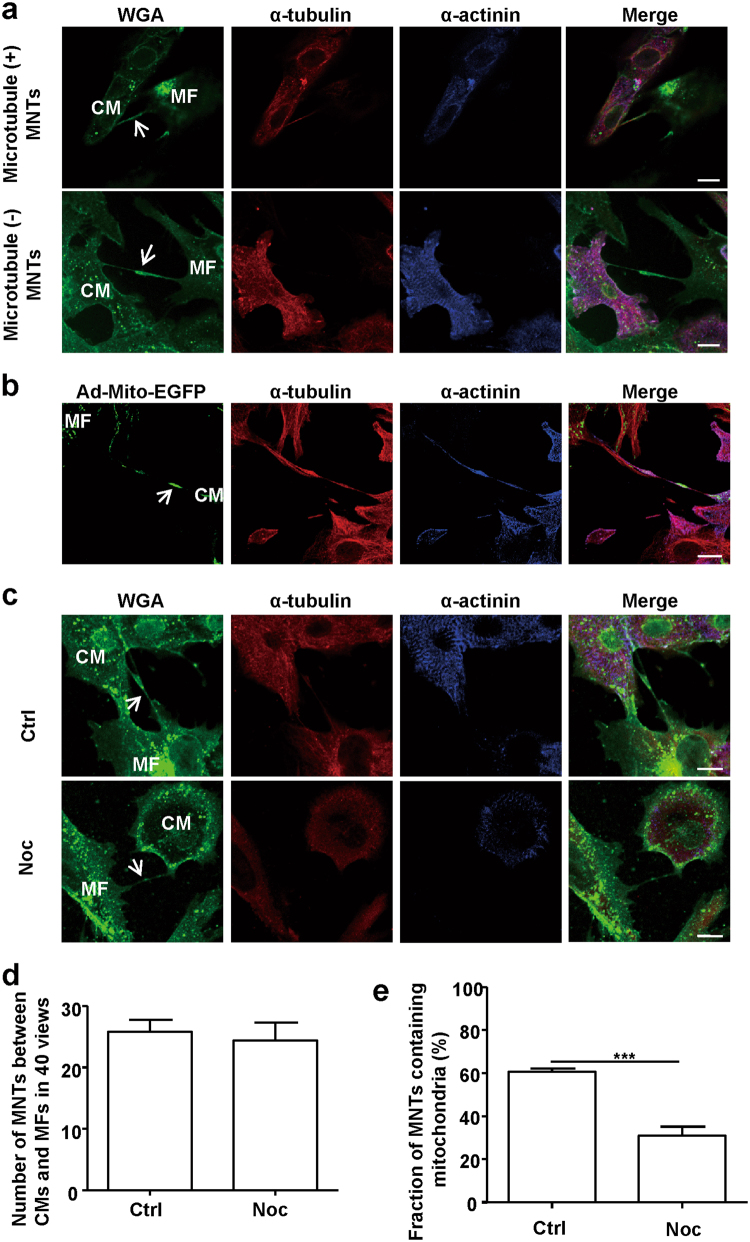
Membrane nanotubes (MNTs) act as "highways" between cells to facilitate the transfer of multiple signals and play an important role in many diseases. Our previous work reported on the transfer of mitochondria via MNTs between cardiomyocytes (CMs) and cardiac myofibroblasts (MFs); however, the elucidation of the underlying mechanism and pathophysiological significance of this transfer requires additional study. In this study, we determined that the mean movement velocity of mitochondria in MNTs between CMs and MFs was approximately 17.5 ± 2.1 nm/s. Meanwhile, treatment with microtubule polymerisation inhibitors nocodazole or colcemid in cell culture decreased mitochondrial velocity, and knockdown of the microtubule motor protein kinesin family member 5B (KIF5B) led to a similar effect, indicating that mitochondrial movement was dependent on microtubules and the motor protein KIF5B. Furthermore, we showed that hypoxia/reoxygenation-induced CM apoptosis was attenuated by coculture with intact or hypoxia/reoxygenation-treated MFs, which transferred mitochondria to CMs. This rescue was prevented either by separating the cells using Transwell culture or by impairing mitochondrial transfer with nocodazole or colcemid treatment. In conclusion, as a novel means of intercellular communication, MNTs rescue distressed CMs from apoptosis by transporting mitochondria along microtubules via KIF5B.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
