

Inflammatory monocytes contribute to the persistence of CXCR3hi CX3CR1lo circulating and lung-resident memory CD8+ T cells following respiratory virus infection
- PMID: 29363162
- PMCID: PMC5916332
- DOI: 10.1111/imcb.12006
Inflammatory monocytes contribute to the persistence of CXCR3hi CX3CR1lo circulating and lung-resident memory CD8+ T cells following respiratory virus infection
Abstract
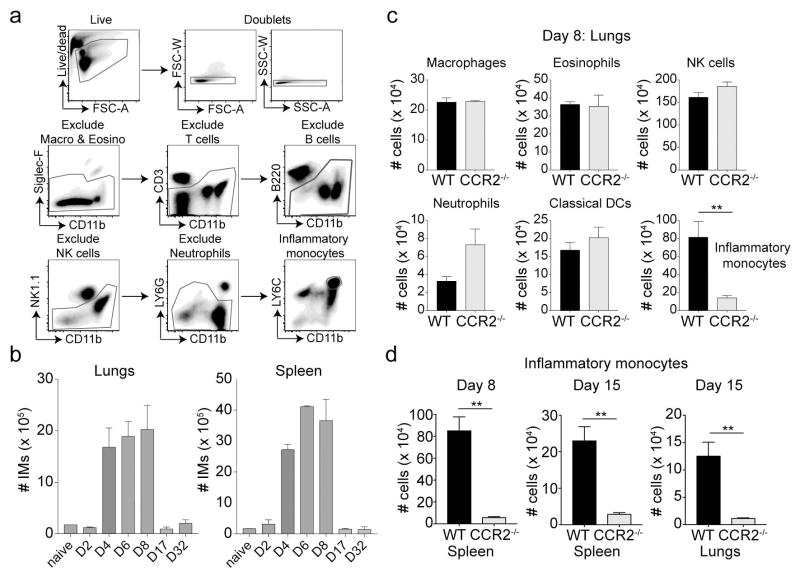
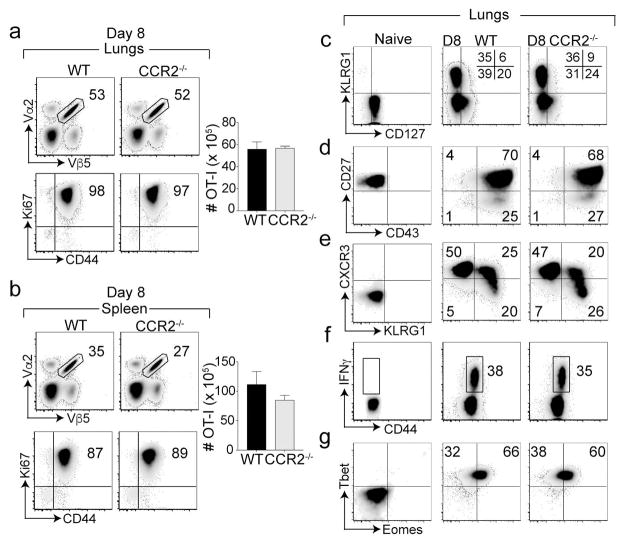
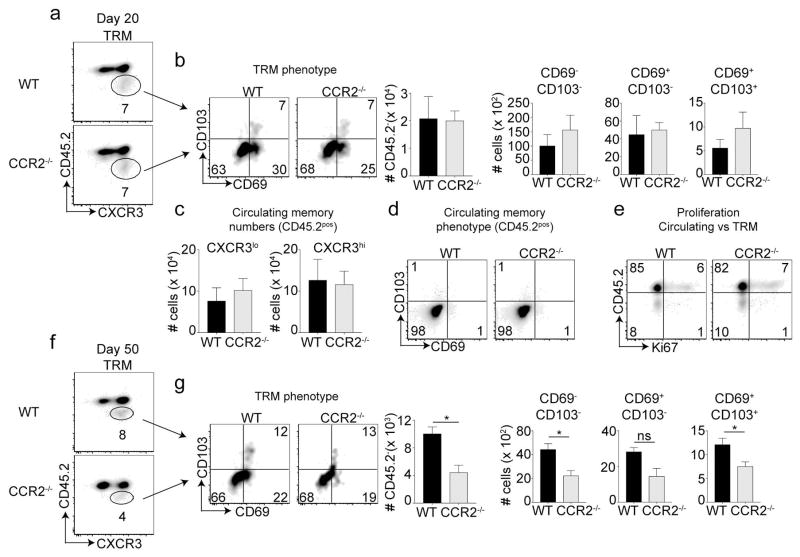
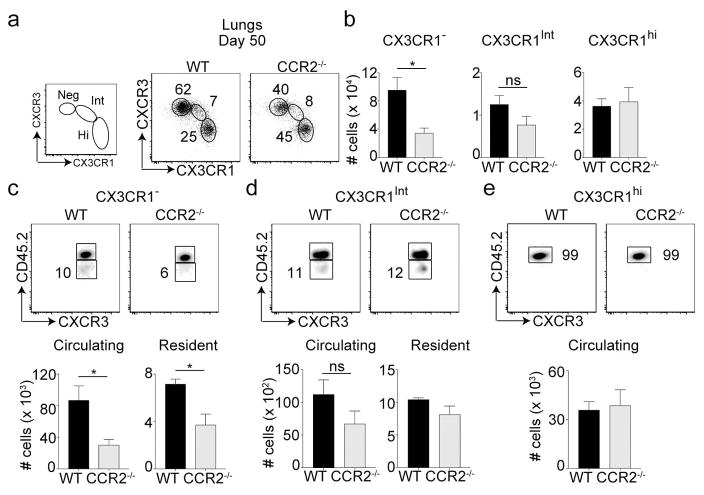
Phenotypically diverse memory CD8+ T cells are present in the lungs that either re-circulate or reside within the tissue. Understanding the key cellular interactions that regulate the generation and then persistence of these different subsets is of great interest. Recently, DNGR-1+ dendritic cell (DC) mediated priming was reported to control the generation of lung-resident but not circulating memory cells following respiratory viral infection. Here, we report an important role for Ly6C+ inflammatory monocytes (IMs) in contributing to the persistence of memory CD8+ T cells but not their generation. Effector CD8+ T cells expanded and contracted normally in the absence of IMs, but the memory compartment declined significantly over time. Quite unexpectedly, this defect was confined to tissue resident and circulating CXCR3hi CX3CR1lo memory cells but not CXCR3hi CX3CR1int and CXCR3lo CX3CR1hi subsets. Thus, two developmentally distinct innate cells orchestrate the generation and persistence of memory T cell subsets following a respiratory virus infection. See also: News and Commentary by Lafouresse & Groom.
Keywords: CD8+ T cells; lung; monocytes; resident memory T cells; vaccinia.
© 2018 Australasian Society for Immunology Inc.
Conflict of interest statement
The authors declare that no conflicts of interest exist related to the work presented herein.
Figures
Comment in
-
Friends help make lasting memories.Immunol Cell Biol. 2018 Apr;96(4):344-346. doi: 10.1111/imcb.12036. Immunol Cell Biol. 2018. PMID: 29687924 No abstract available.
Similar articles
-
Lack of B Lymphocytes Enhances CD8 T Cell-Mediated Resistance against Respiratory Viral Infection but Compromises Memory Cell Formation.J Virol. 2020 Jan 17;94(3):e01877-19. doi: 10.1128/JVI.01877-19. Print 2020 Jan 17. J Virol. 2020. PMID: 31723023 Free PMC article.
-
GITR differentially affects lung effector T cell subpopulations during influenza virus infection.J Leukoc Biol. 2020 Jun;107(6):953-970. doi: 10.1002/JLB.4AB1219-254R. Epub 2020 Mar 3. J Leukoc Biol. 2020. PMID: 32125017
-
A Distinct Lung-Interstitium-Resident Memory CD8(+) T Cell Subset Confers Enhanced Protection to Lower Respiratory Tract Infection.Cell Rep. 2016 Aug 16;16(7):1800-9. doi: 10.1016/j.celrep.2016.07.037. Epub 2016 Aug 4. Cell Rep. 2016. PMID: 27498869 Free PMC article.
-
Persistence in Temporary Lung Niches: A Survival Strategy of Lung-Resident Memory CD8+ T Cells.Viral Immunol. 2017 Jul/Aug;30(6):438-450. doi: 10.1089/vim.2017.0016. Epub 2017 Apr 18. Viral Immunol. 2017. PMID: 28418771 Free PMC article. Review.
-
Genealogy, Dendritic Cell Priming, and Differentiation of Tissue-Resident Memory CD8+ T Cells.Front Immunol. 2018 Jul 31;9:1751. doi: 10.3389/fimmu.2018.01751. eCollection 2018. Front Immunol. 2018. PMID: 30108585 Free PMC article. Review.
Cited by
-
Divergence of Tissue-Memory T Cells: Distribution and Function-Based Classification.Cold Spring Harb Perspect Biol. 2020 Oct 1;12(10):a037762. doi: 10.1101/cshperspect.a037762. Cold Spring Harb Perspect Biol. 2020. PMID: 32816841 Free PMC article. Review.
-
Tissue resident memory T cells in the respiratory tract.Mucosal Immunol. 2022 Mar;15(3):379-388. doi: 10.1038/s41385-021-00461-z. Epub 2021 Oct 20. Mucosal Immunol. 2022. PMID: 34671115 Free PMC article. Review.
-
Poxviridae Pneumonia.Adv Exp Med Biol. 2024;1451:183-204. doi: 10.1007/978-3-031-57165-7_12. Adv Exp Med Biol. 2024. PMID: 38801579 Review.
-
Alterations in CX3CL1 Levels and Its Role in Viral Pathogenesis.Int J Mol Sci. 2024 Apr 18;25(8):4451. doi: 10.3390/ijms25084451. Int J Mol Sci. 2024. PMID: 38674036 Free PMC article. Review.
-
The Memory T Cell "Communication Web" in Context with Gastrointestinal Disorders-How Memory T Cells Affect Their Surroundings and How They Are Influenced by It.Cells. 2022 Sep 6;11(18):2780. doi: 10.3390/cells11182780. Cells. 2022. PMID: 36139354 Free PMC article. Review.
References
-
- Kohlmeier JE, Woodland DL. Immunity to respiratory viruses. Annu Rev Immunol. 2009;27:61–82. - PubMed
-
- Woodland DL, Kohlmeier JE. Migration, maintenance and recall of memory T cells in peripheral tissues. Nat Rev Immunol. 2009;9:153–161. - PubMed
-
- Rutishauser RL, Kaech SM. Generating diversity: transcriptional regulation of effector and memory CD8 T-cell differentiation. Immunol Rev. 2010;235:219–233. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
