

Life under the Microscope: Single-Molecule Fluorescence Highlights the RNA World
- PMID: 29363314
- PMCID: PMC5918467
- DOI: 10.1021/acs.chemrev.7b00519
Life under the Microscope: Single-Molecule Fluorescence Highlights the RNA World
Abstract
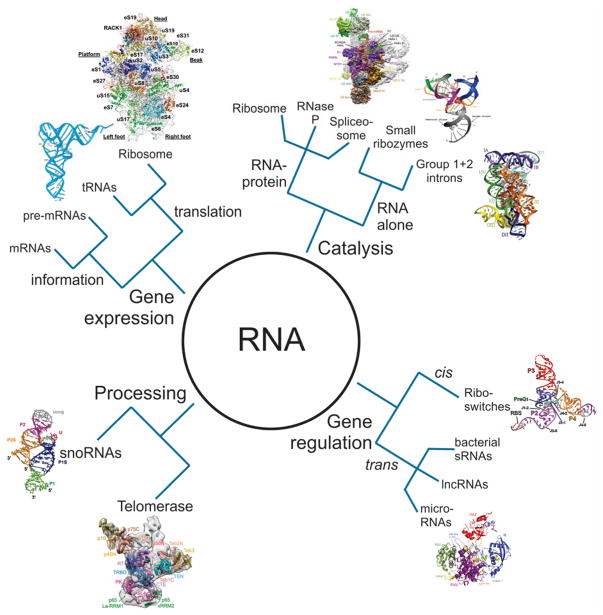
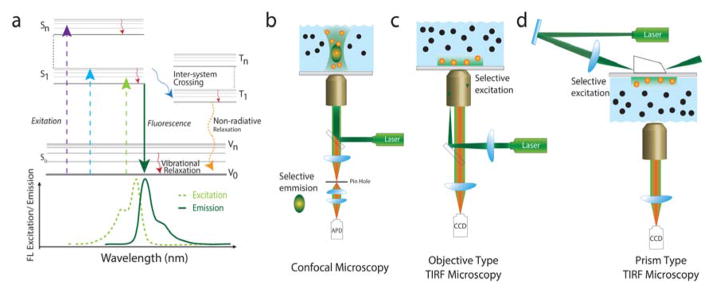
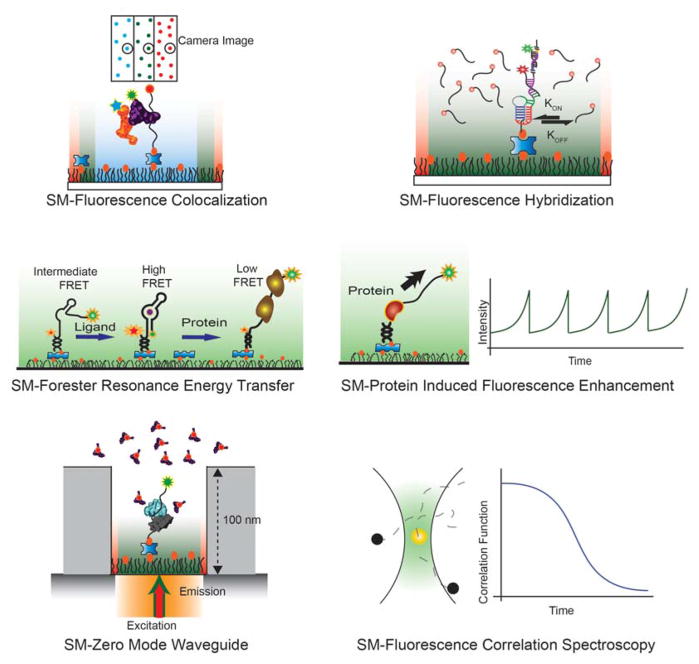
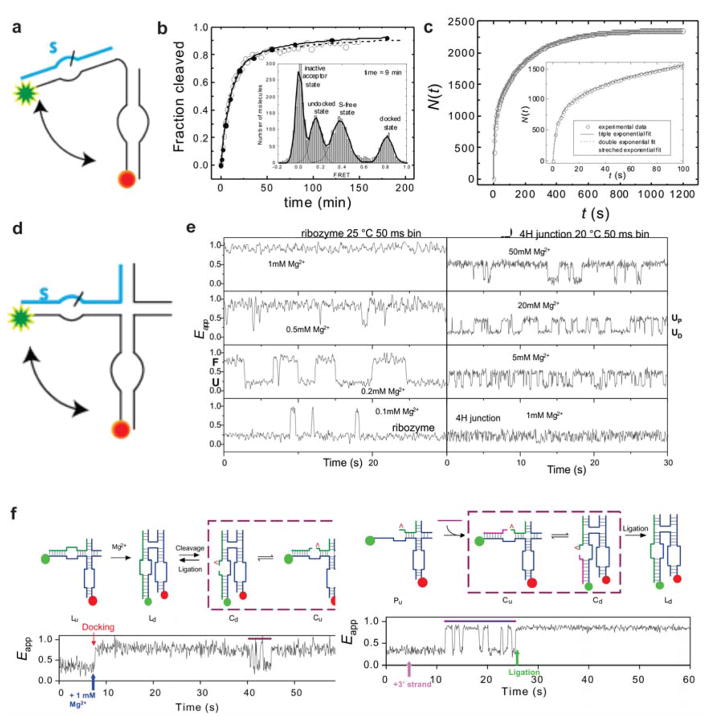
The emergence of single-molecule (SM) fluorescence techniques has opened up a vast new toolbox for exploring the molecular basis of life. The ability to monitor individual biomolecules in real time enables complex, dynamic folding pathways to be interrogated without the averaging effect of ensemble measurements. In parallel, modern biology has been revolutionized by our emerging understanding of the many functions of RNA. In this comprehensive review, we survey SM fluorescence approaches and discuss how the application of these tools to RNA and RNA-containing macromolecular complexes in vitro has yielded significant insights into the underlying biology. Topics covered include the three-dimensional folding landscapes of a plethora of isolated RNA molecules, their assembly and interactions in RNA-protein complexes, and the relation of these properties to their biological functions. In all of these examples, the use of SM fluorescence methods has revealed critical information beyond the reach of ensemble averages.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
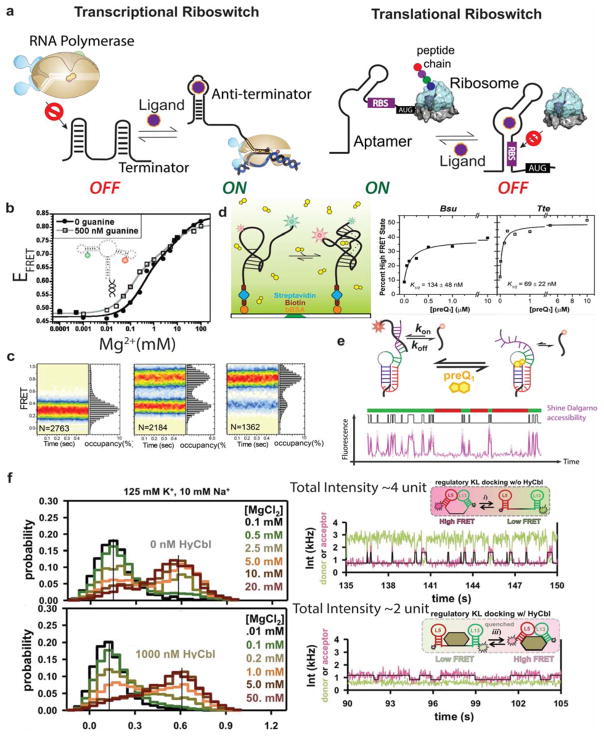
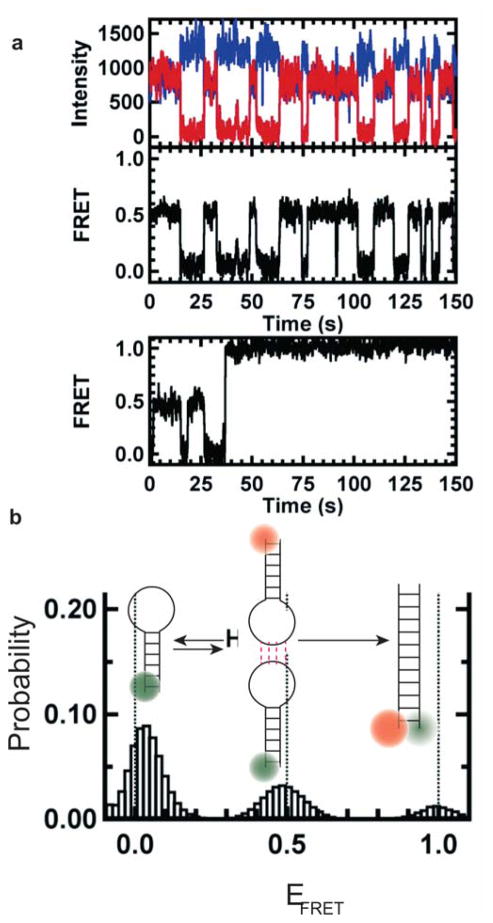
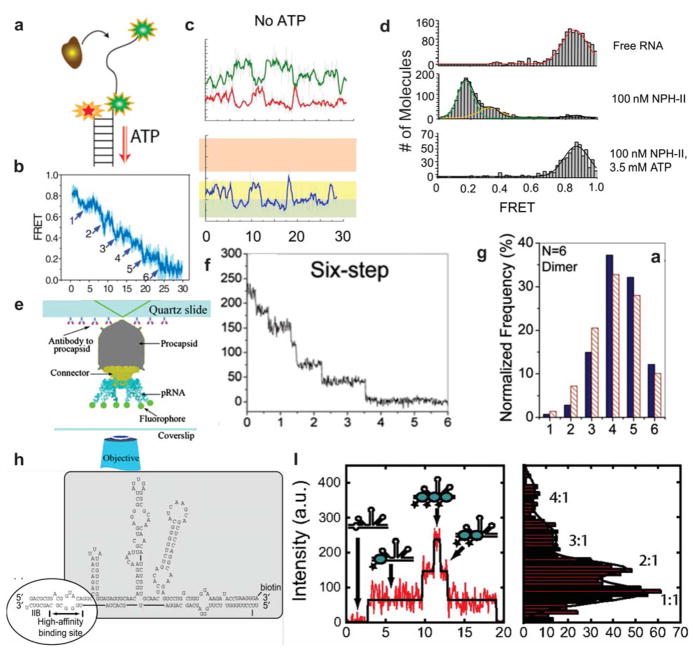
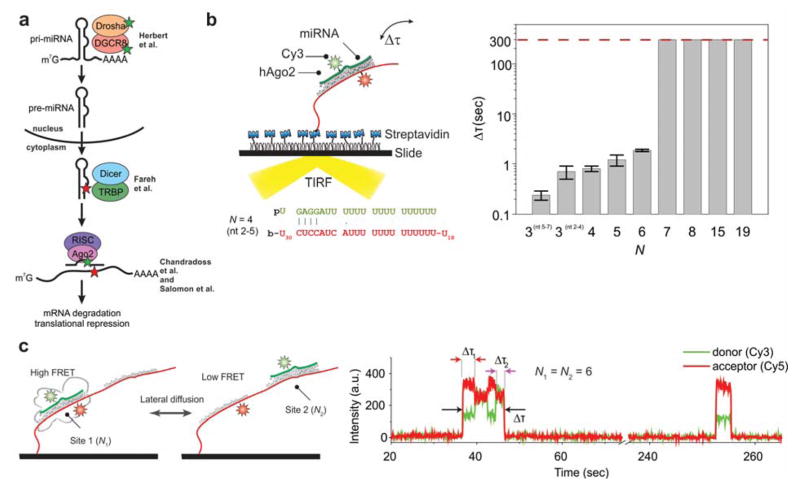
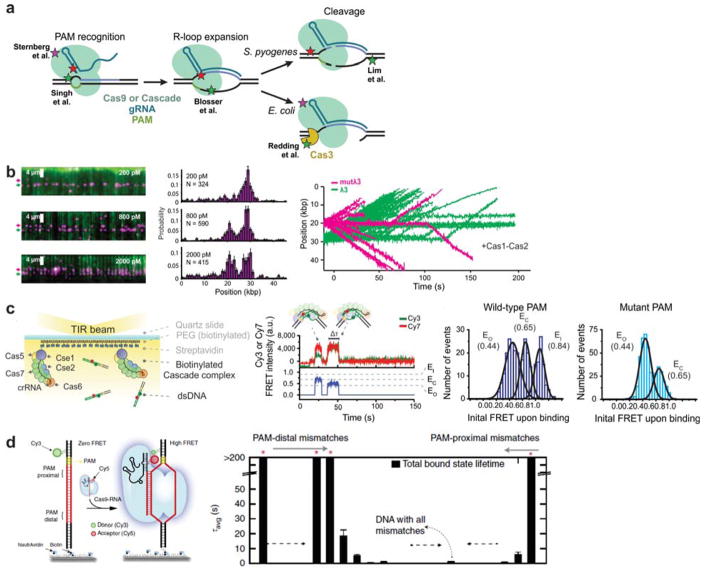
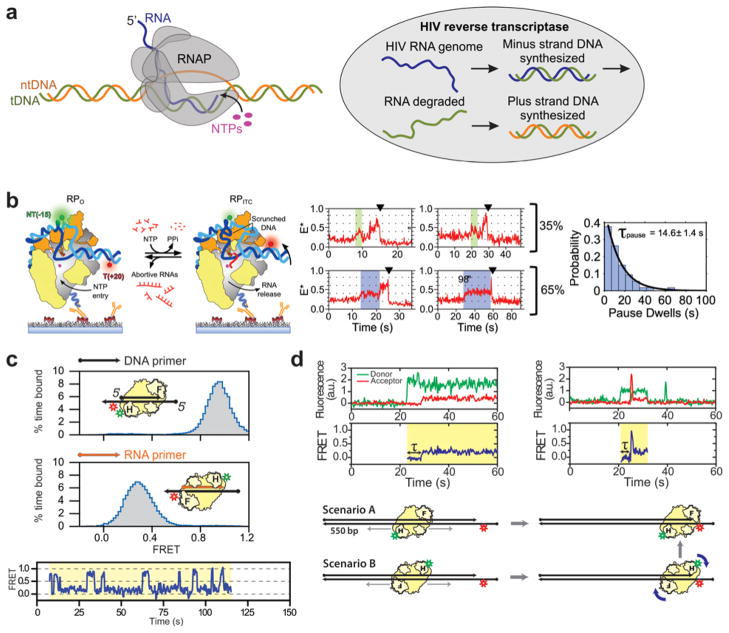
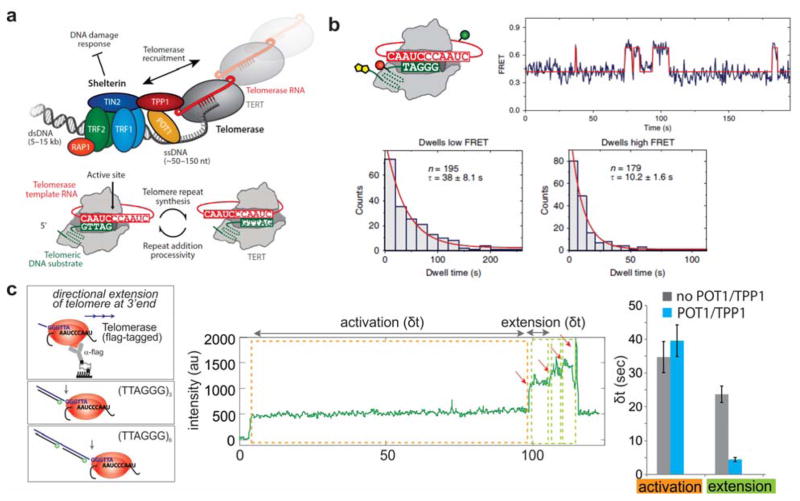
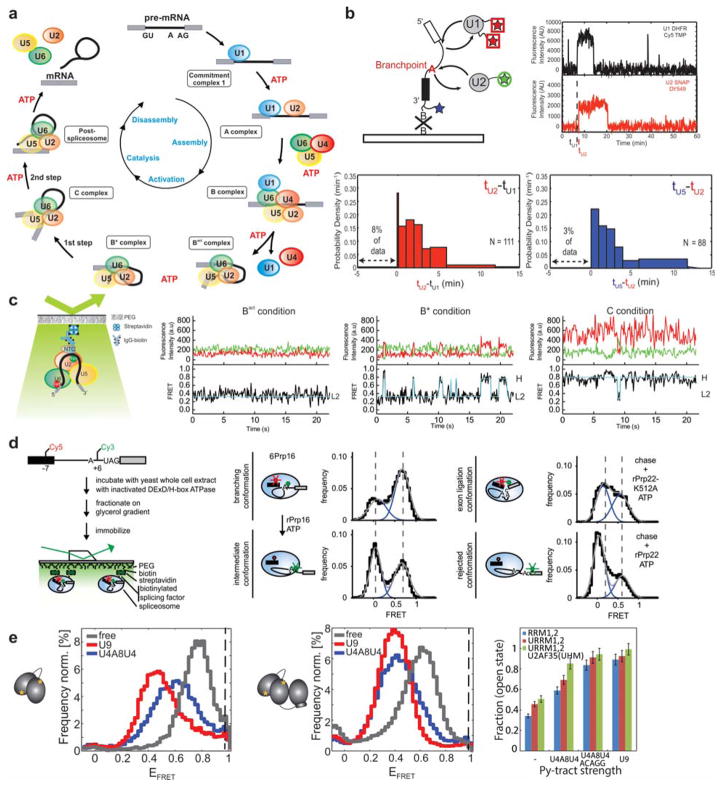
References
-
- Kruger K, Grabowski PJ, Zaug AJ, Sands J, Gottschling DE, Cech TR. Self-Splicing Rna: Autoexcision and Autocyclization of the Ribosomal Rna Intervening Sequence of Tetrahymena. Cell. 1982;31:147–157. - PubMed
-
- Guerrier-Takada C, Gardiner K, Marsh T, Pace N, Altman S. The Rna Moiety of Ribonuclease P Is the Catalytic Subunit of the Enzyme. Cell. 1983;35:849–857. - PubMed
-
- Moerner WE, Kador L. Optical Detection and Spectroscopy of Single Molecules in a Solid. Phys Rev Lett. 1989;62:2535–2538. - PubMed
-
- Carninci P, Kasukawa T, Katayama S, Gough J, Frith MC, Maeda N, Oyama R, Ravasi T, Lenhard B, Wells C, et al. The Transcriptional Landscape of the Mammalian Genome. Science. 2005;309:1559–1563. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
