

Nitrate modulates stem cell dynamics in Arabidopsis shoot meristems through cytokinins
- PMID: 29363596
- PMCID: PMC5819446
- DOI: 10.1073/pnas.1718670115
Nitrate modulates stem cell dynamics in Arabidopsis shoot meristems through cytokinins
Abstract
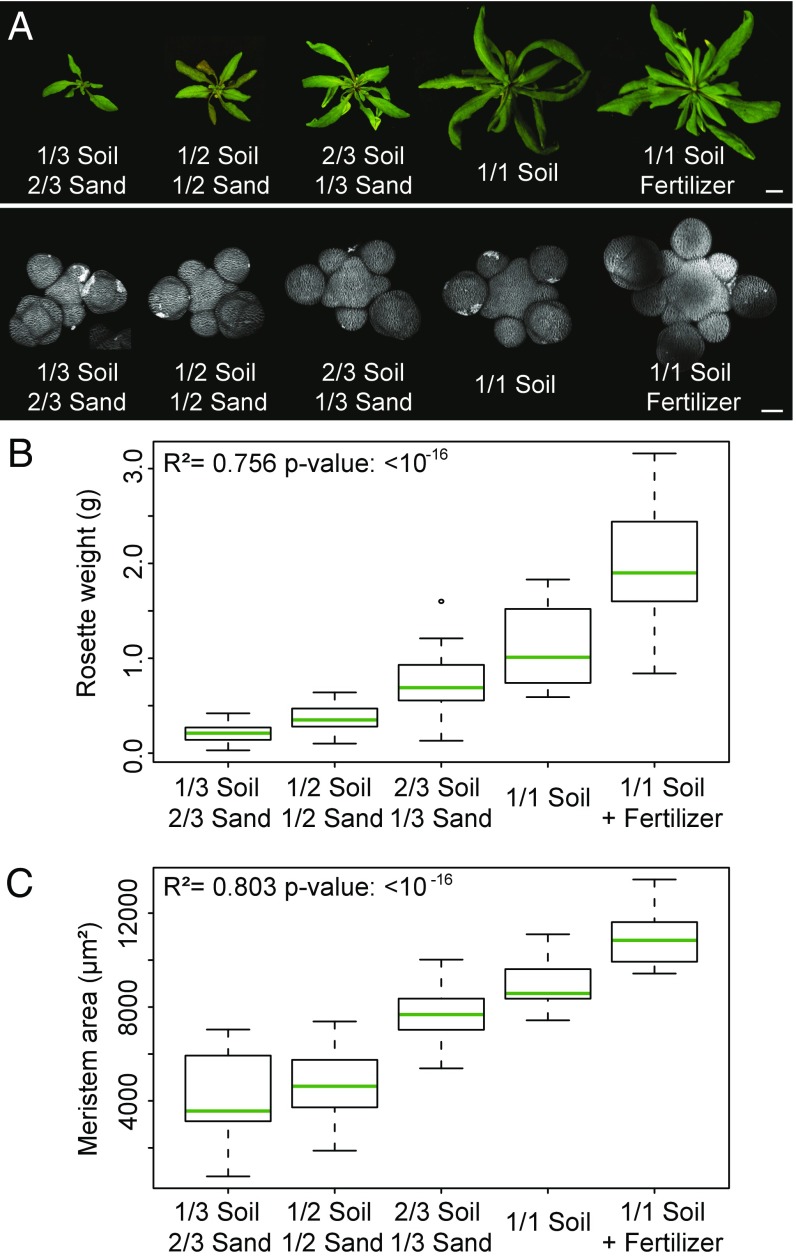
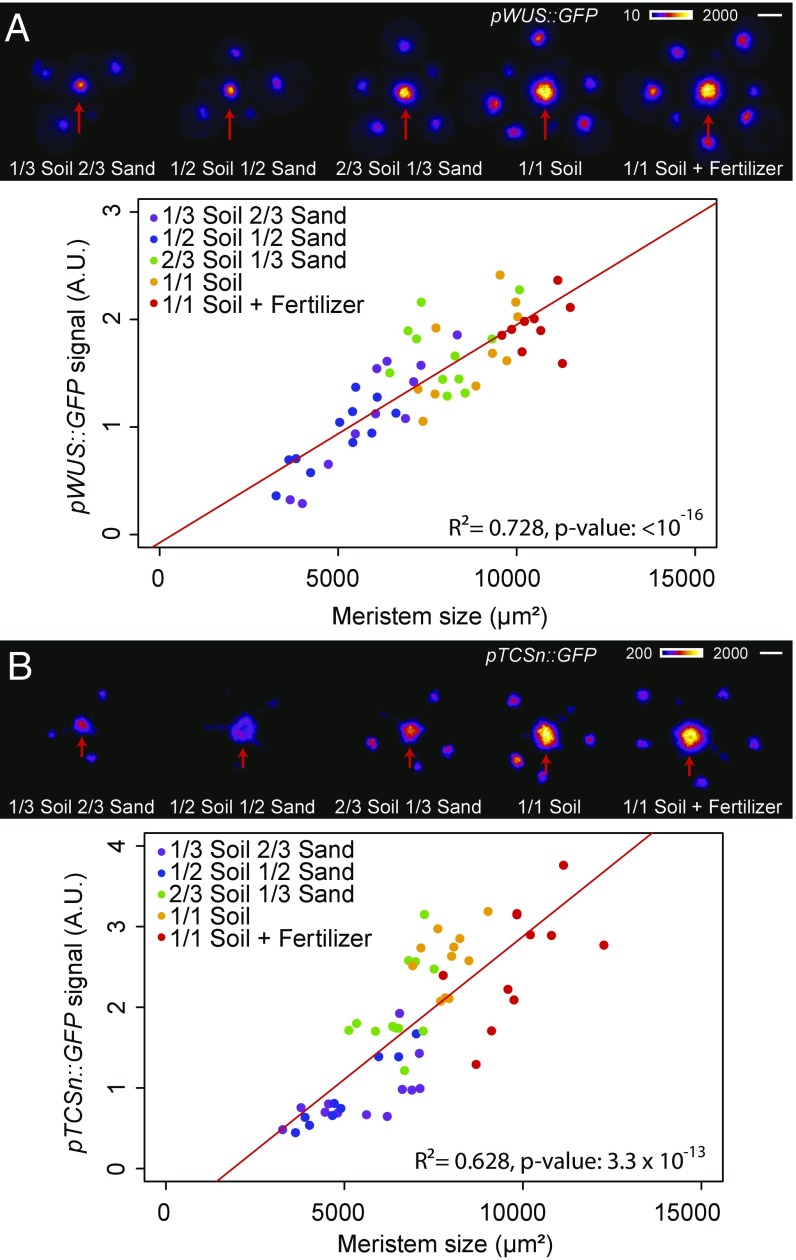
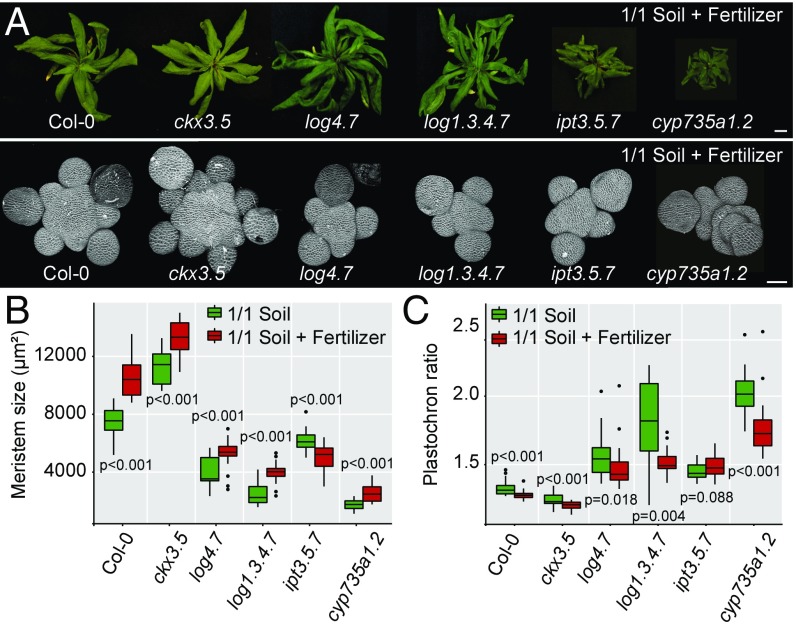
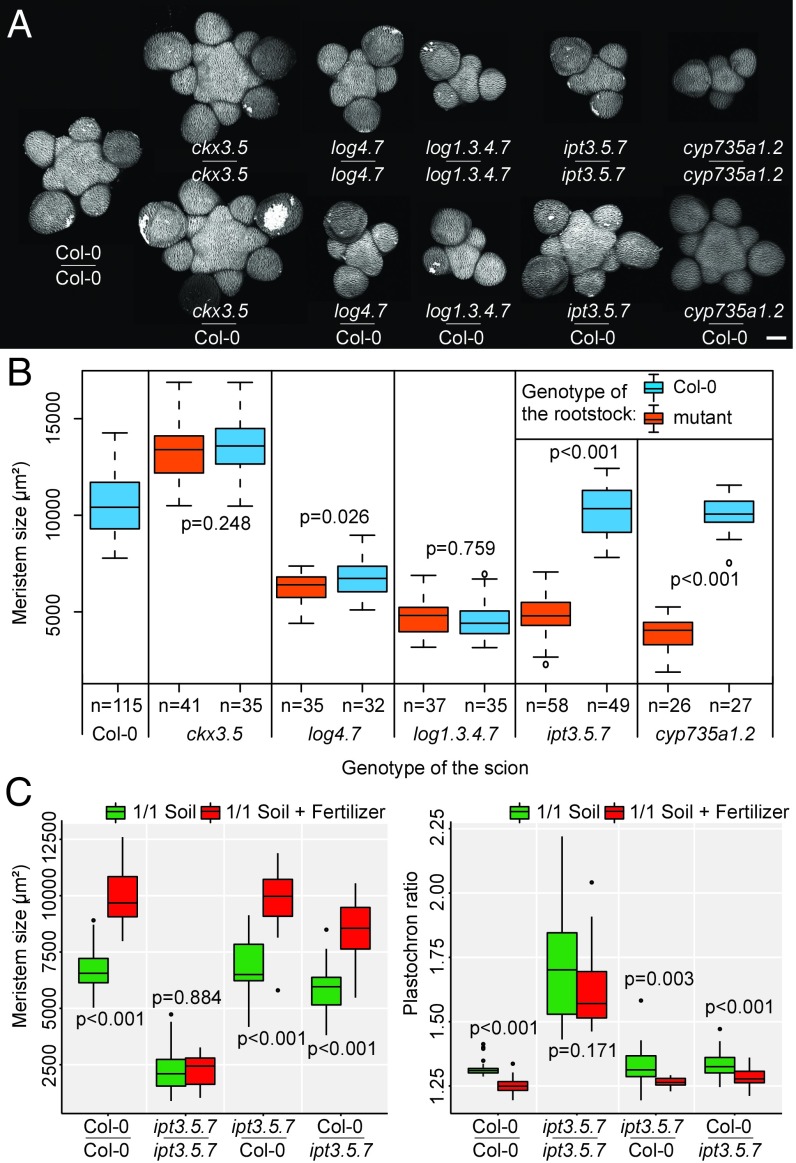
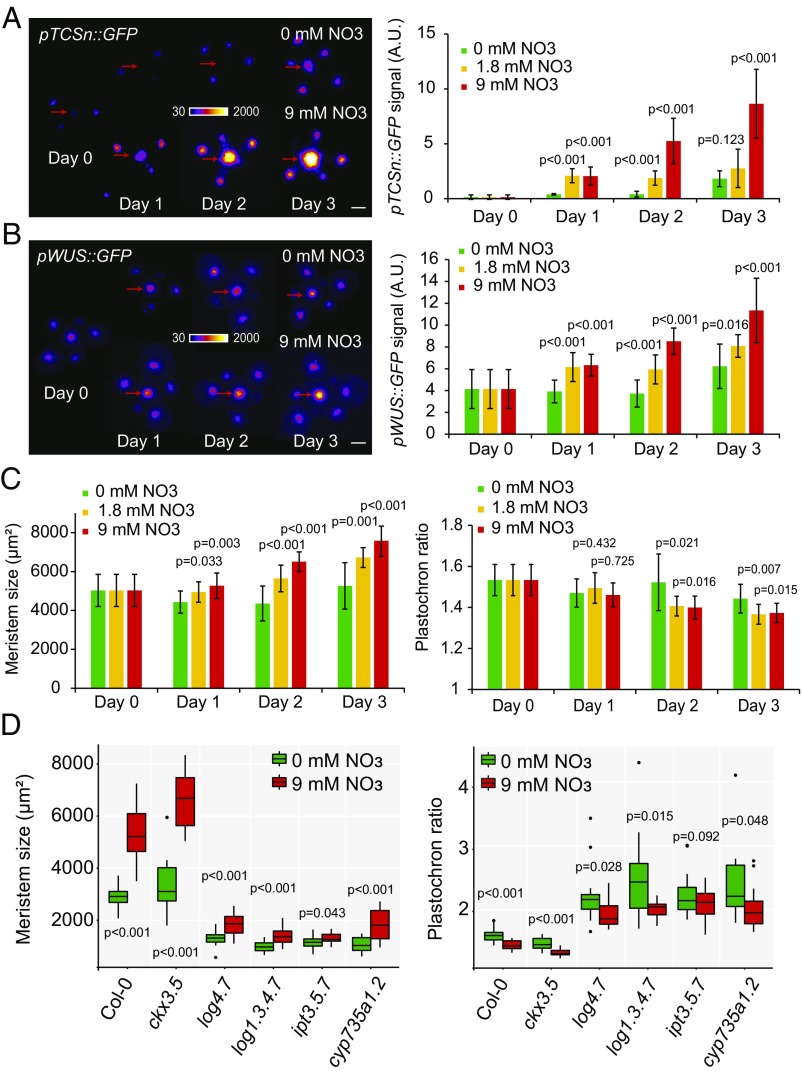
The shoot apical meristem (SAM) is responsible for the generation of all the aerial parts of plants. Given its critical role, dynamical changes in SAM activity should play a central role in the adaptation of plant architecture to the environment. Using quantitative microscopy, grafting experiments, and genetic perturbations, we connect the plant environment to the SAM by describing the molecular mechanism by which cytokinins signal the level of nutrient availability to the SAM. We show that a systemic signal of cytokinin precursors mediates the adaptation of SAM size and organogenesis rate to the availability of mineral nutrients by modulating the expression of WUSCHEL, a key regulator of stem cell homeostasis. In time-lapse experiments, we further show that this mechanism allows meristems to adapt to rapid changes in nitrate concentration, and thereby modulate their rate of organ production to the availability of mineral nutrients within a few days. Our work sheds light on the role of the stem cell regulatory network by showing that it not only maintains meristem homeostasis but also allows plants to adapt to rapid changes in the environment.
Keywords: Arabidopsis; cytokinin hormones; plant development; plant nutrition; shoot apical meristem.
Copyright © 2018 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Comment in
-
Nitrate, NO and ROS Signaling in Stem Cell Homeostasis.Trends Plant Sci. 2018 Dec;23(12):1041-1044. doi: 10.1016/j.tplants.2018.09.010. Epub 2018 Oct 10. Trends Plant Sci. 2018. PMID: 30316685
References
-
- Krouk G, et al. A framework integrating plant growth with hormones and nutrients. Trends Plant Sci. 2011;16:178–182. - PubMed
-
- Werner T, Schmülling T. Cytokinin action in plant development. Curr Opin Plant Biol. 2009;12:527–538. - PubMed
-
- Rahayu YS, et al. Root-derived cytokinins as long-distance signals for NO3–Induced stimulation of leaf growth. J Exp Bot. 2005;56:1143–1152. - PubMed
-
- Walch-Liu P, Neumann G, Bangerth F, Engels C. Rapid effects of nitrogen form on leaf morphogenesis in tobacco. J Exp Bot. 2000;51:227–237. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
