

Once and only once: mechanisms of centriole duplication and their deregulation in disease
- PMID: 29363672
- PMCID: PMC5969912
- DOI: 10.1038/nrm.2017.127
Once and only once: mechanisms of centriole duplication and their deregulation in disease
Abstract
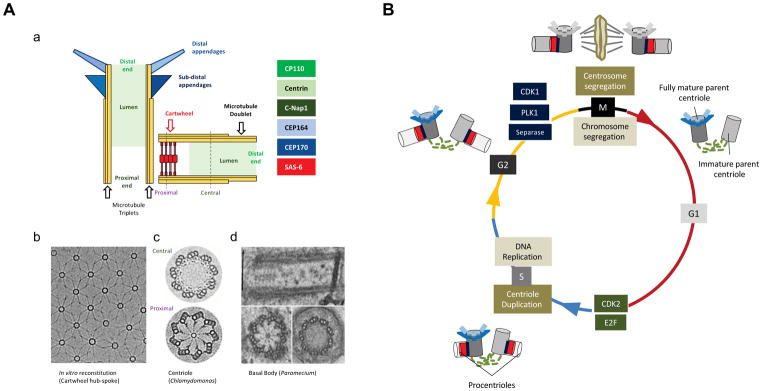
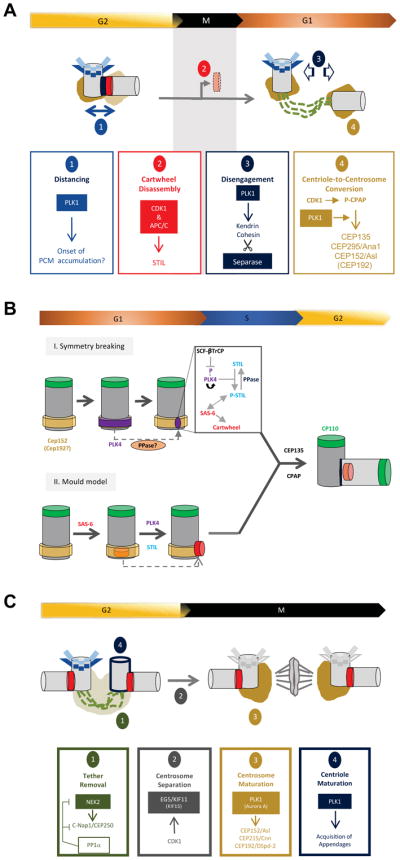
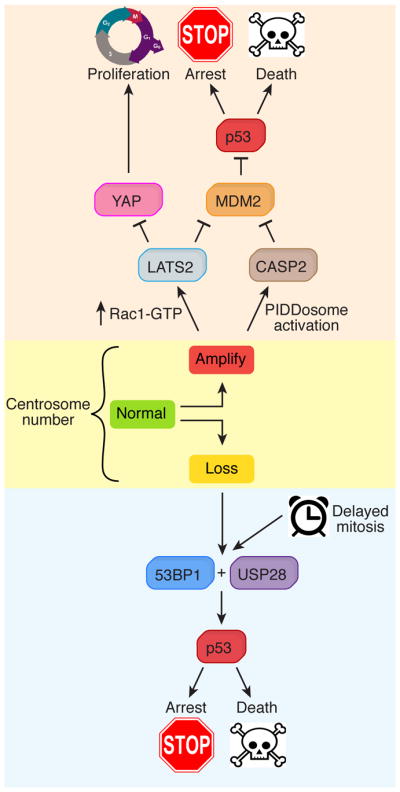
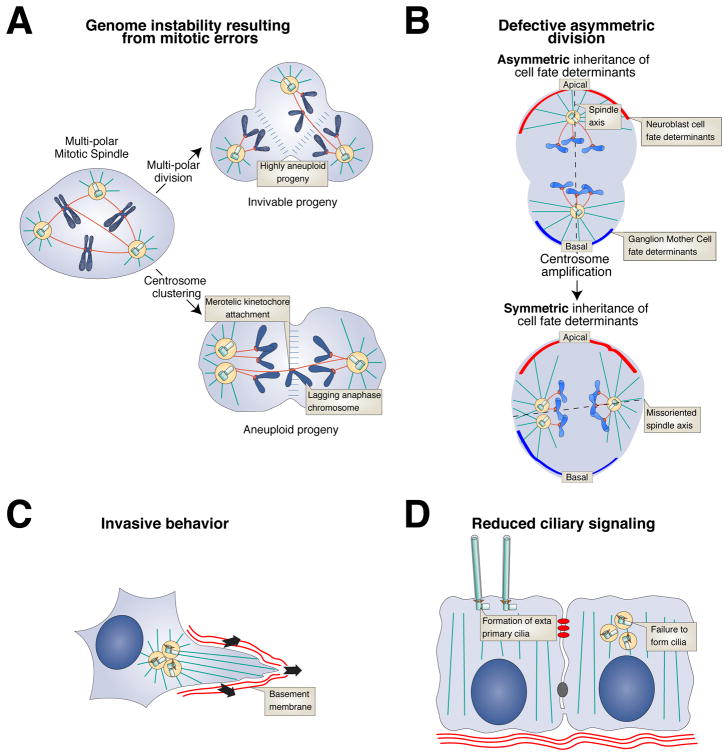
Centrioles are conserved microtubule-based organelles that form the core of the centrosome and act as templates for the formation of cilia and flagella. Centrioles have important roles in most microtubule-related processes, including motility, cell division and cell signalling. To coordinate these diverse cellular processes, centriole number must be tightly controlled. In cycling cells, one new centriole is formed next to each pre-existing centriole in every cell cycle. Advances in imaging, proteomics, structural biology and genome editing have revealed new insights into centriole biogenesis, how centriole numbers are controlled and how alterations in these processes contribute to diseases such as cancer and neurodevelopmental disorders. Moreover, recent work has uncovered the existence of surveillance pathways that limit the proliferation of cells with numerical centriole aberrations. Owing to this progress, we now have a better understanding of the molecular mechanisms governing centriole biogenesis, opening up new possibilities for targeting these pathways in the context of human disease.
Figures
References
-
- Nigg EA, Raff JW. Centrioles, centrosomes, and cilia in health and disease. Cell. 2009;139:663–678. - PubMed
-
- Bornens M. The centrosome in cells and organisms. Science. 2012;335:422–426. - PubMed
-
- Conduit PT, Wainman A, Raff JW. Centrosome function and assembly in animal cells. Nat Rev Mol Cell Biol. 2015;16:611–624. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
