

An essential thioredoxin is involved in the control of the cell cycle in the bacterium Caulobacter crescentus
- PMID: 29367337
- PMCID: PMC5846133
- DOI: 10.1074/jbc.RA117.001042
An essential thioredoxin is involved in the control of the cell cycle in the bacterium Caulobacter crescentus
Abstract
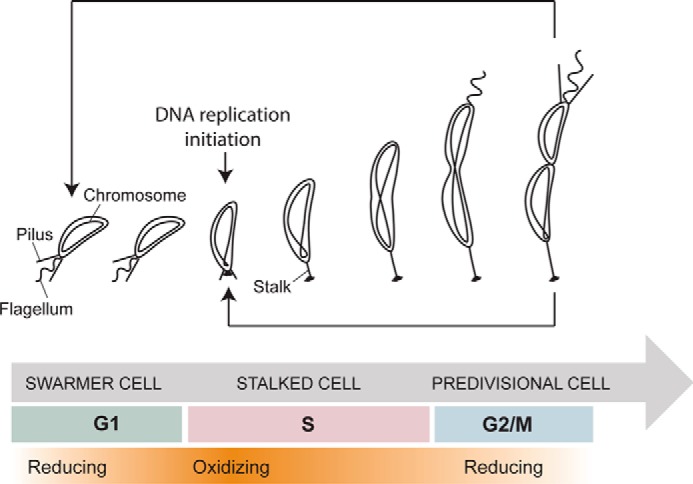
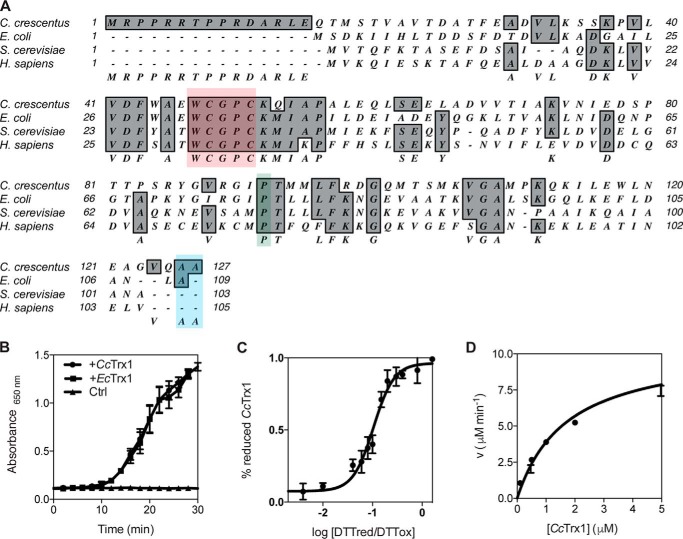
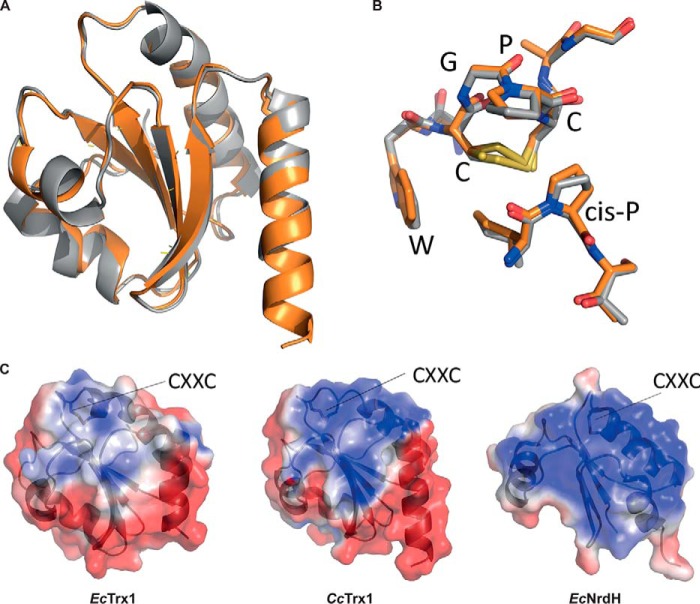
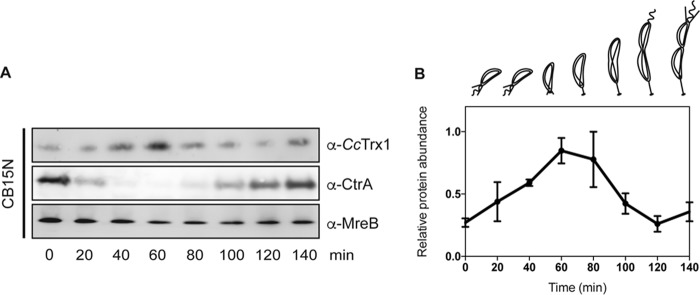
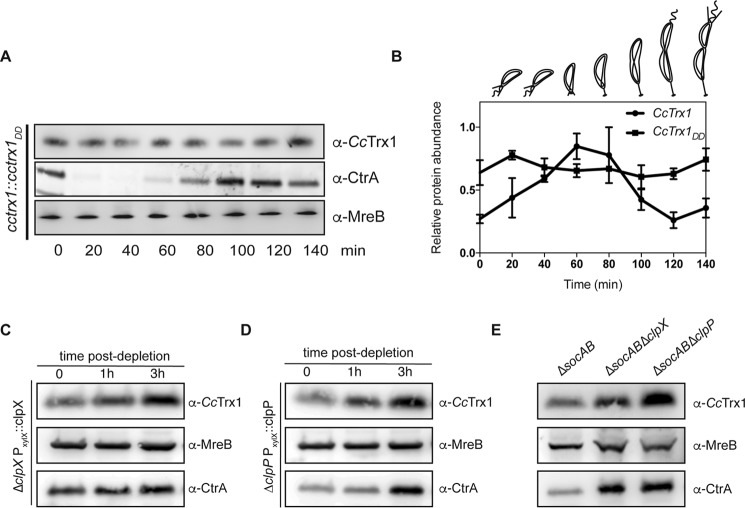
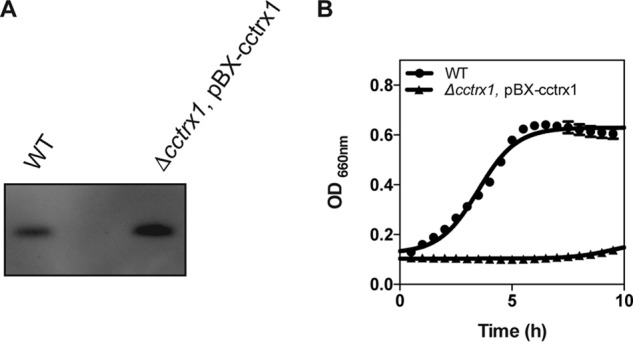
Thioredoxins (Trxs) are antioxidant proteins that are conserved among all species. These proteins have been extensively studied and perform reducing reactions on a broad range of substrates. Here, we identified Caulobacter crescentus Trx1 (CCNA_03653; CcTrx1) as an oxidoreductase that is involved in the cell cycle progression of this model bacterium and is required to sustain life. Intriguingly, the abundance of CcTrx1 varies throughout the C. crescentus cell cycle: although the expression of CcTrx1 is induced in stalked cells, right before DNA replication initiation, CcTrx1 is actively degraded by the ClpXP protease in predivisional cells. Importantly, we demonstrated that regulation of the abundance of CcTrx1 is crucial for cell growth and survival as modulating CcTrx1 levels leads to cell death. Finally, we also report a comprehensive biochemical and structural characterization of this unique and essential Trx. The requirement to precisely control the abundance of CcTrx1 for cell survival underlines the importance of redox control for optimal cell cycle progression in C. crescentus.
Keywords: Caulobacter crescentus; antioxidant; bacteria; crystal structure; oxidation-reduction (redox); redox regulation; thioredoxin.
© 2018 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Laurent T. C., Moore E. C., and Reichard P. (1964) Enzymatic synthesis of deoxyribonucleotides. IV. Isolation and characterization of thioredoxin, the hydrogen donor from Escherichia coli B. J. Biol. Chem. 239, 3436–3444 - PubMed
-
- Moore E. C., Reichard P., and Thelander L. (1964) Enzymatic synthesis of deoxyribonucleotides. V. Purification and properties of thioredoxin reductase from Escherichia coli B. J. Biol. Chem. 239, 3445–3452 - PubMed
-
- Arts I. S., Vertommen D., Baldin F., Laloux G., and Collet J. F. (2016) Comprehensively characterizing the thioredoxin interactome in vivo highlights the central role played by this ubiquitous oxidoreductase in redox control. Mol. Cell. Proteomics 15, 2125–2140 10.1074/mcp.M115.056440 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
