

The life of U6 small nuclear RNA, from cradle to grave
- PMID: 29367453
- PMCID: PMC5855946
- DOI: 10.1261/rna.065136.117
The life of U6 small nuclear RNA, from cradle to grave
Abstract
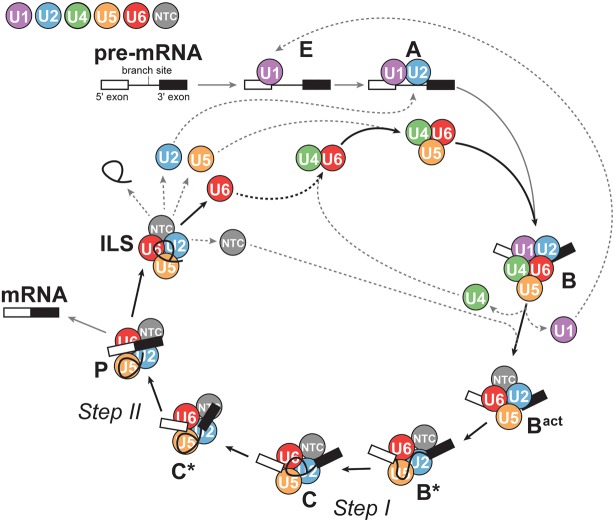
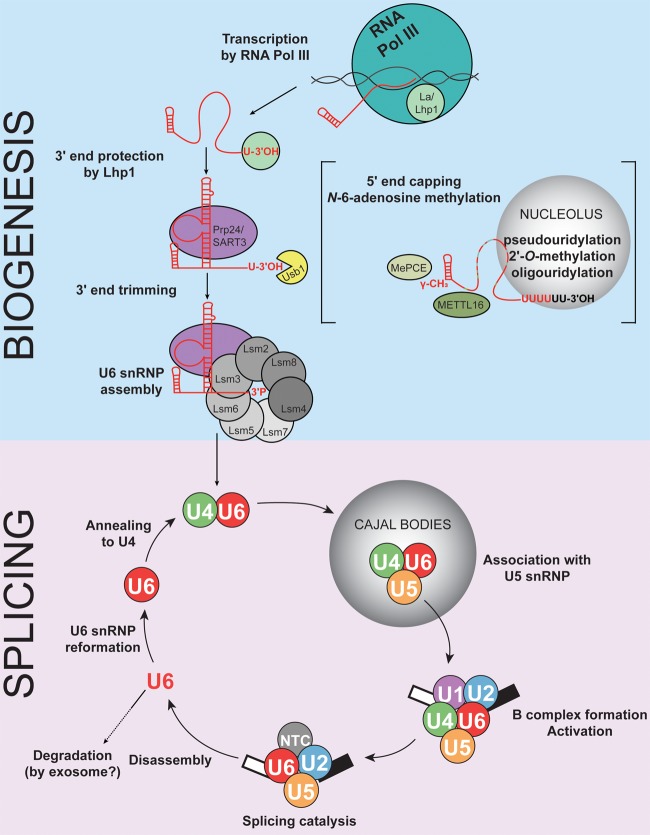
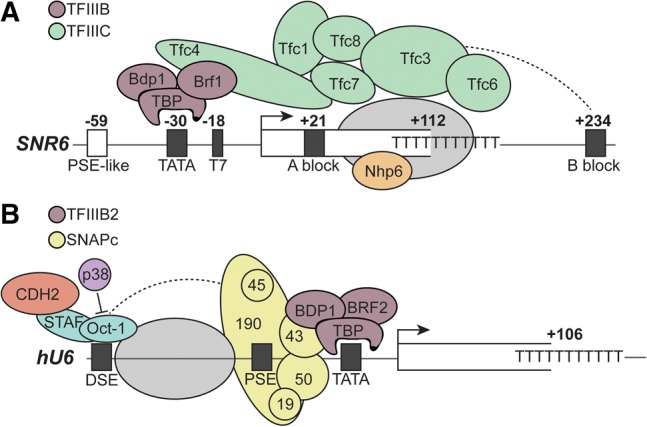
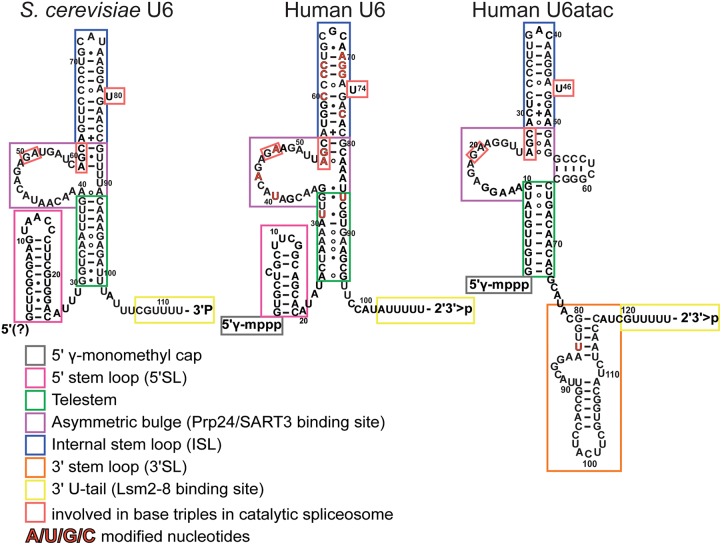
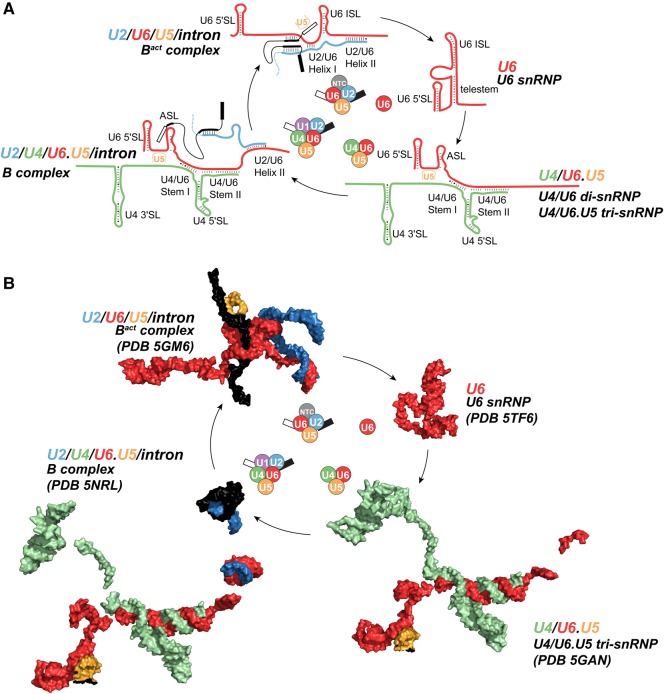
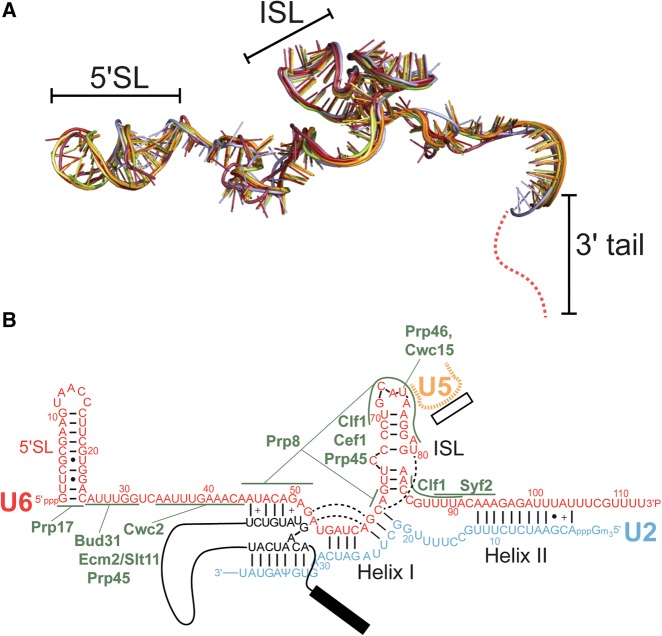
Removal of introns from precursor messenger RNA (pre-mRNA) and some noncoding transcripts is an essential step in eukaryotic gene expression. In the nucleus, this process of RNA splicing is carried out by the spliceosome, a multi-megaDalton macromolecular machine whose core components are conserved from yeast to humans. In addition to many proteins, the spliceosome contains five uridine-rich small nuclear RNAs (snRNAs) that undergo an elaborate series of conformational changes to correctly recognize the splice sites and catalyze intron removal. Decades of biochemical and genetic data, along with recent cryo-EM structures, unequivocally demonstrate that U6 snRNA forms much of the catalytic core of the spliceosome and is highly dynamic, interacting with three snRNAs, the pre-mRNA substrate, and >25 protein partners throughout the splicing cycle. This review summarizes the current state of knowledge on how U6 snRNA is synthesized, modified, incorporated into snRNPs and spliceosomes, recycled, and degraded.
Keywords: U6 gene transcription; U6 snRNA; U6 snRNP; spliceosome.
© 2018 Didychuk et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures
References
-
- Afroz T, Cienikova Z, Cléry A, Allain FH. 2015. One, two, three, four! How multiple RRMs read the genome sequence. Methods Enzymol 558: 235–278. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases