

T cells specific for post-translational modifications escape intrathymic tolerance induction
- PMID: 29367624
- PMCID: PMC5783942
- DOI: 10.1038/s41467-017-02763-y
T cells specific for post-translational modifications escape intrathymic tolerance induction
Abstract
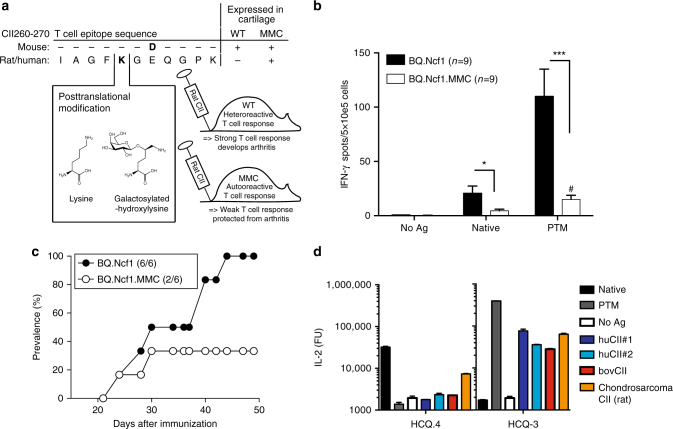
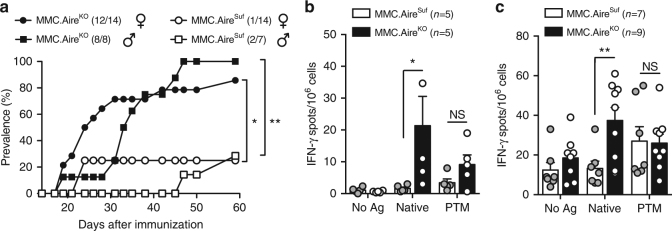
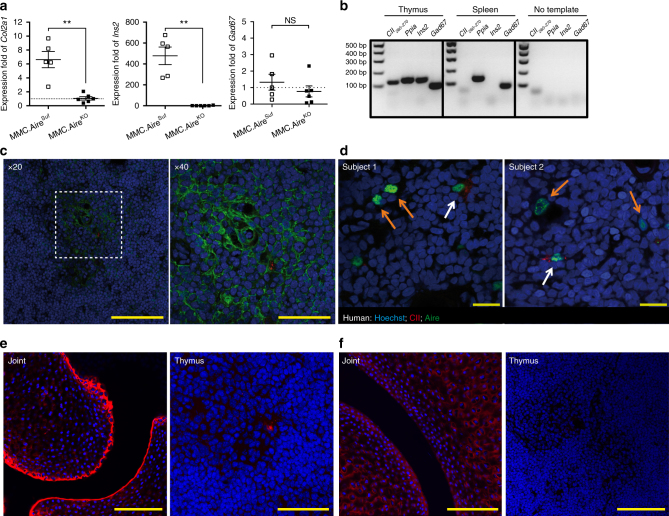
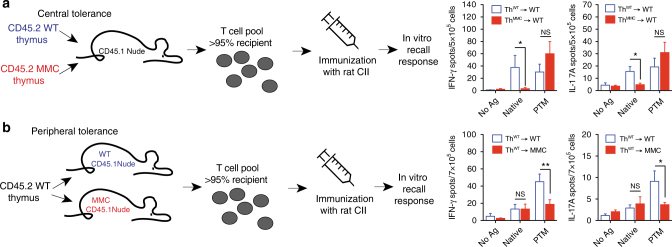
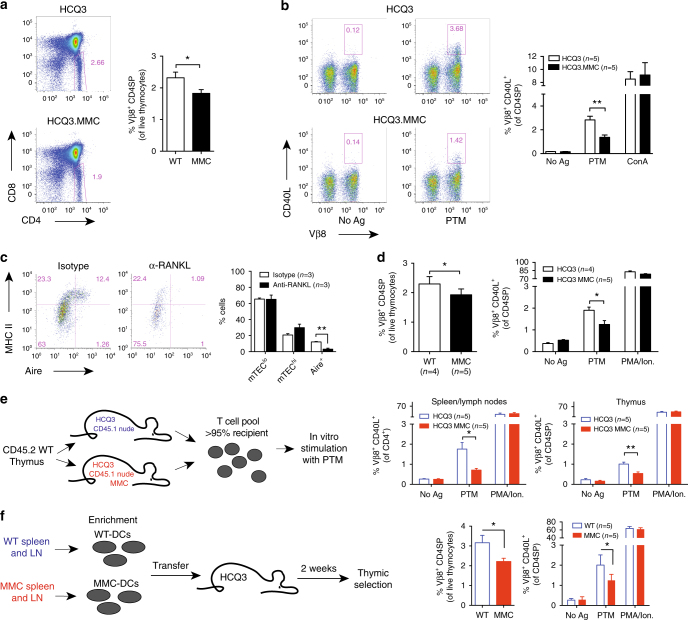
Establishing effective central tolerance requires the promiscuous expression of tissue-restricted antigens by medullary thymic epithelial cells. However, whether central tolerance also extends to post-translationally modified proteins is not clear. Here we show a mouse model of autoimmunity in which disease development is dependent on post-translational modification (PTM) of the tissue-restricted self-antigen collagen type II. T cells specific for the non-modified antigen undergo efficient central tolerance. By contrast, PTM-reactive T cells escape thymic selection, though the PTM variant constitutes the dominant form in the periphery. This finding implies that the PTM protein is absent in the thymus, or present at concentrations insufficient to induce negative selection of developing thymocytes and explains the lower level of tolerance induction against the PTM antigen. As the majority of self-antigens are post-translationally modified, these data raise the possibility that T cells specific for other self-antigens naturally subjected to PTM may escape central tolerance induction by a similar mechanism.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Klein L, Hinterberger M, Wirnsberger G, Kyewski B. Antigen presentation in the thymus for positive selection and central tolerance induction. Nat. Publ. Group. 2009;9:833–844. - PubMed
-
- Derbinski, J. & Kyewski, B. How thymic antigen presenting cells sample the body′s self-antigens. Curr. Opin. Immunol. 22, 592-600 (2010). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
