

Diversity of meso-scale architecture in human and non-human connectomes
- PMID: 29367627
- PMCID: PMC5783945
- DOI: 10.1038/s41467-017-02681-z
Diversity of meso-scale architecture in human and non-human connectomes
Abstract
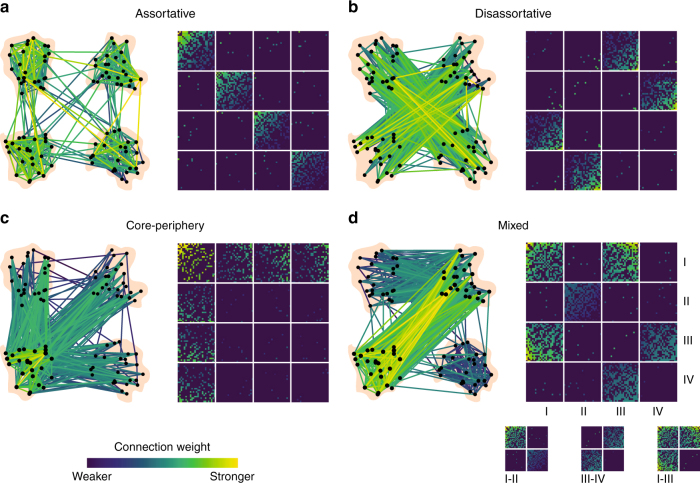
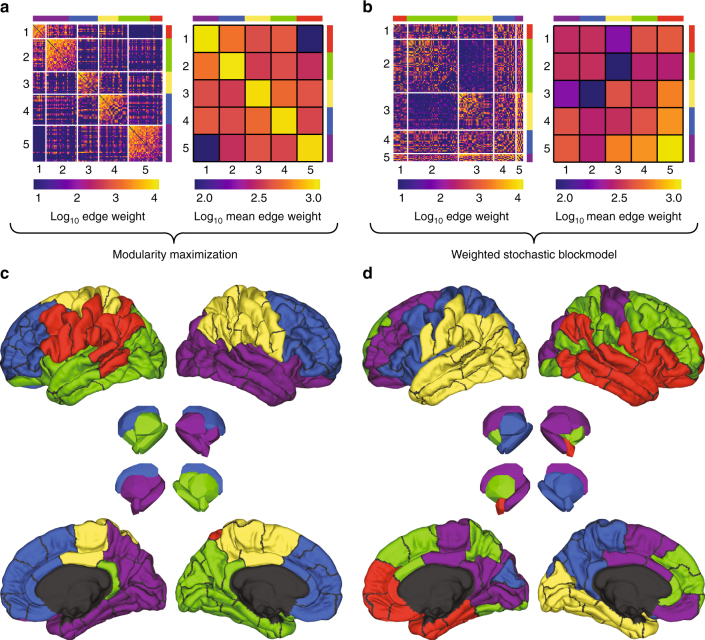
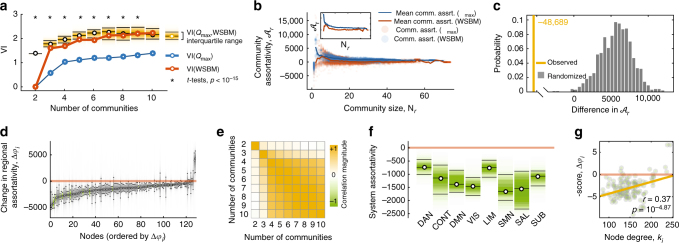
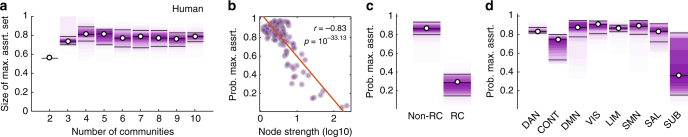
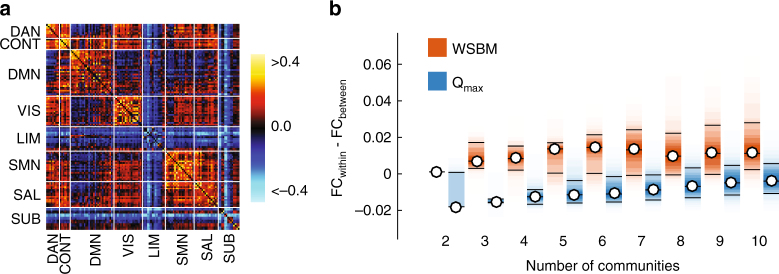
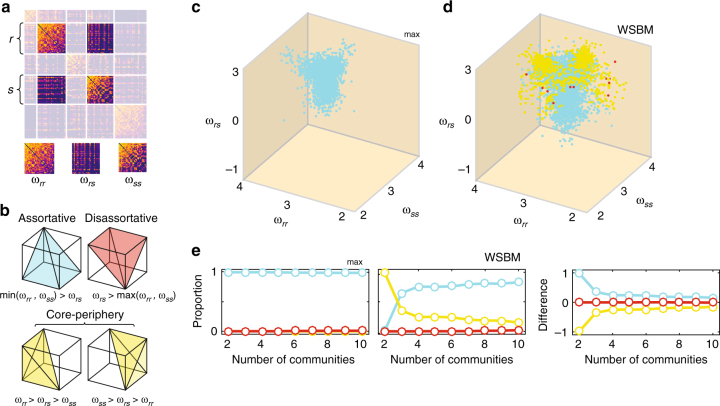
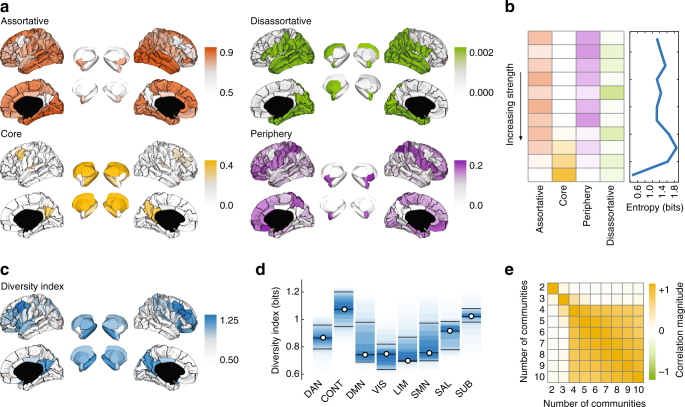
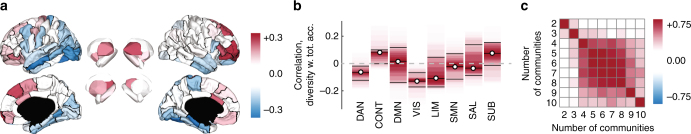
Brain function is reflected in connectome community structure. The dominant view is that communities are assortative and segregated from one another, supporting specialized information processing. However, this view precludes the possibility of non-assortative communities whose complex inter-community interactions could engender a richer functional repertoire. We use weighted stochastic blockmodels to uncover the meso-scale architecture of Drosophila, mouse, rat, macaque, and human connectomes. We find that most communities are assortative, though others form core-periphery and disassortative structures, which better recapitulate observed patterns of functional connectivity and gene co-expression in human and mouse connectomes compared to standard community detection techniques. We define measures for quantifying the diversity of communities in which brain regions participate, showing that this measure is peaked in control and subcortical systems in humans, and that inter-individual differences are correlated with cognitive performance. Our report paints a more diverse portrait of connectome communities and demonstrates their cognitive relevance.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
The Connectomes: Methods of White Matter Tractography and Contributions of Resting State fMRI.Semin Ultrasound CT MR. 2021 Oct;42(5):507-522. doi: 10.1053/j.sult.2021.07.007. Epub 2021 Aug 1. Semin Ultrasound CT MR. 2021. PMID: 34537118
-
Constraints and spandrels of interareal connectomes.Nat Commun. 2016 Dec 7;7:13812. doi: 10.1038/ncomms13812. Nat Commun. 2016. PMID: 27924867 Free PMC article.
-
An edge-centric perspective on the human connectome: link communities in the brain.Philos Trans R Soc Lond B Biol Sci. 2014 Oct 5;369(1653):20130527. doi: 10.1098/rstb.2013.0527. Philos Trans R Soc Lond B Biol Sci. 2014. PMID: 25180305 Free PMC article.
-
The meso-connectomes of mouse, marmoset, and macaque: network organization and the emergence of higher cognition.Cereb Cortex. 2024 May 2;34(5):bhae174. doi: 10.1093/cercor/bhae174. Cereb Cortex. 2024. PMID: 38771244 Free PMC article. Review.
-
White matter pathways and social cognition.Neurosci Biobehav Rev. 2018 Jul;90:350-370. doi: 10.1016/j.neubiorev.2018.04.015. Epub 2018 Apr 20. Neurosci Biobehav Rev. 2018. PMID: 29684403 Free PMC article. Review.
Cited by
-
Determining the Hierarchical Architecture of the Human Brain Using Subject-Level Clustering of Functional Networks.Sci Rep. 2019 Dec 17;9(1):19290. doi: 10.1038/s41598-019-55738-y. Sci Rep. 2019. PMID: 31848397 Free PMC article.
-
A Transcriptome Community-and-Module Approach of the Human Mesoconnectome.Entropy (Basel). 2021 Aug 11;23(8):1031. doi: 10.3390/e23081031. Entropy (Basel). 2021. PMID: 34441171 Free PMC article.
-
Path-dependent connectivity, not modularity, consistently predicts controllability of structural brain networks.Netw Neurosci. 2020 Nov 1;4(4):1091-1121. doi: 10.1162/netn_a_00157. eCollection 2020. Netw Neurosci. 2020. PMID: 33195950 Free PMC article.
-
Homological landscape of human brain functional sub-circuits.bioRxiv [Preprint]. 2023 Dec 22:2023.12.22.573062. doi: 10.1101/2023.12.22.573062. bioRxiv. 2023. Update in: Mathematics (Basel). 2024 Feb;12(3). doi: 10.3390/math12030455. PMID: 38187668 Free PMC article. Updated. Preprint.
-
Deep phenotypic analysis of psychiatric features in genetically defined cohorts: application to XYY syndrome.J Neurodev Disord. 2023 Feb 20;15(1):8. doi: 10.1186/s11689-023-09476-y. J Neurodev Disord. 2023. PMID: 36803654 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
