

Palisade cell shape affects the light-induced chloroplast movements and leaf photosynthesis
- PMID: 29367686
- PMCID: PMC5784166
- DOI: 10.1038/s41598-018-19896-9
Palisade cell shape affects the light-induced chloroplast movements and leaf photosynthesis
Abstract
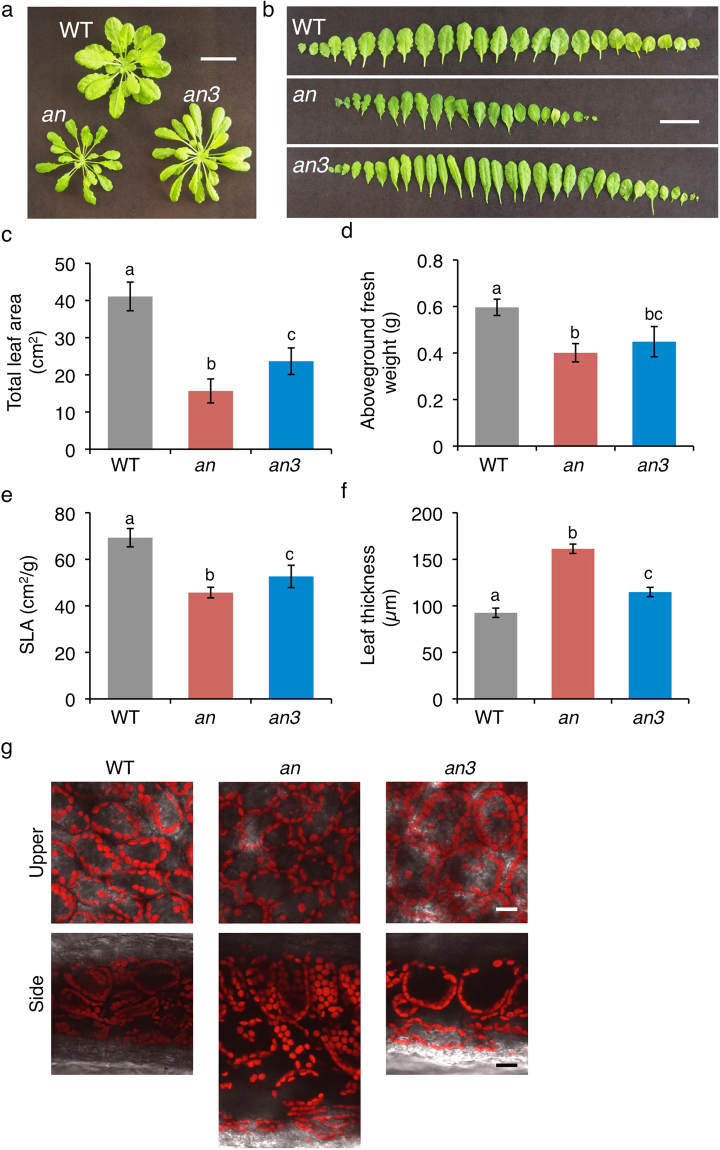
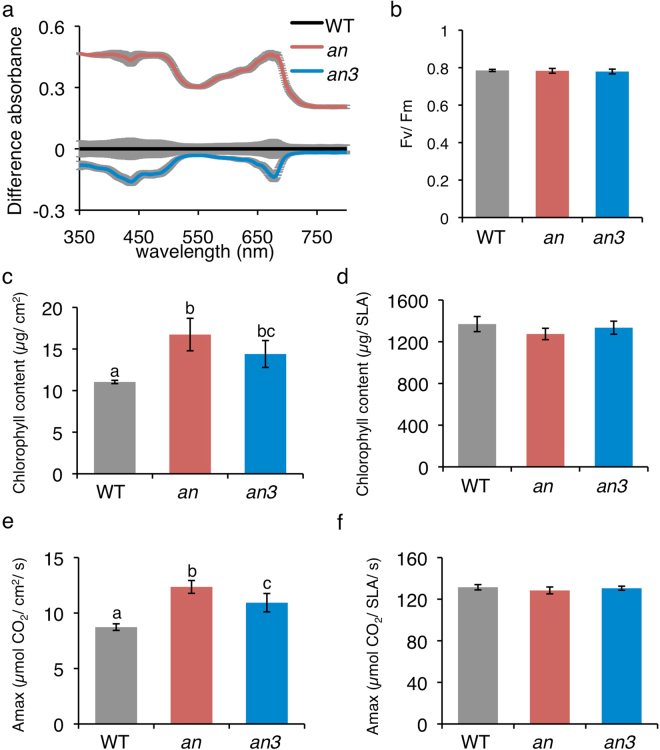
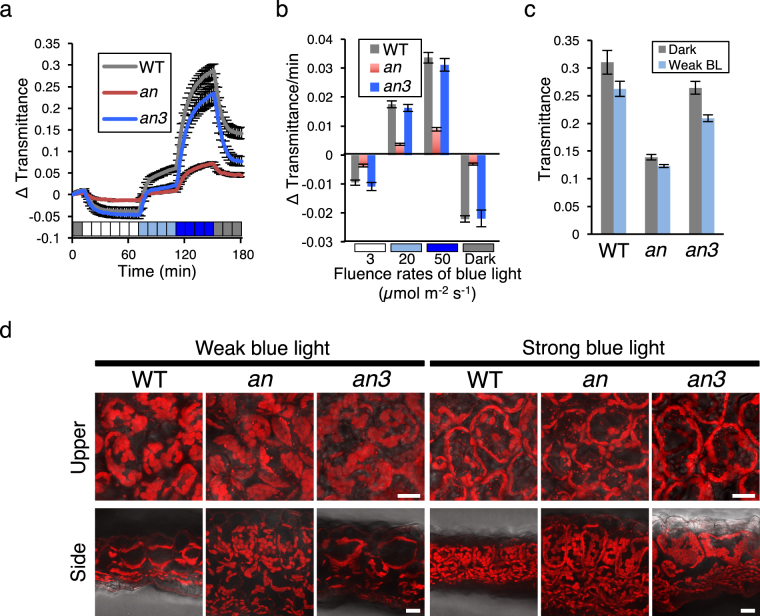
Leaf photosynthesis is regulated by multiple factors that help the plant to adapt to fluctuating light conditions. Leaves of sun-light-grown plants are thicker and contain more columnar palisade cells than those of shade-grown plants. Light-induced chloroplast movements are also essential for efficient leaf photosynthesis and facilitate efficient light utilization in leaf cells. Previous studies have demonstrated that leaves of most of the sun-grown plants exhibited no or very weak chloroplast movements and could accomplish efficient photosynthesis under strong light. To examine the relationship between palisade cell shape, chloroplast movement and distribution, and leaf photosynthesis, we used an Arabidopsis thaliana mutant, angustifolia (an), which has thick leaves that contain columnar palisade cells similar to those in the sun-grown plants. In the highly columnar cells of an mutant leaves, chloroplast movements were restricted. Nevertheless, under white light condition (at 120 µmol m-2 s-1), the an mutant plants showed higher chlorophyll content per unit leaf area and, thus, higher light absorption by the leaves than the wild type, which resulted in enhanced photosynthesis per unit leaf area. Our findings indicate that coordinated regulation of leaf cell shape and chloroplast movement according to the light conditions is pivotal for efficient leaf photosynthesis.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Suetsugu, N. & Wada, M. Chloroplast photorelocation movement: A sophisticated strategy for chloroplasts to perform efficient photosynthesis in Advances in photosynthesis-Fundamental aspects (ed. Najafpour, M. M.) 215–234 (InTech, 2012).
-
- Terashima I, Hikosaka K. Comparative ecophysiology of leaf and canopy photosynthesis. Plant Cell Environ. 1995;18:1111–1128. doi: 10.1111/j.1365-3040.1995.tb00623.x. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
