

Inhibition of EV71 by curcumin in intestinal epithelial cells
- PMID: 29370243
- PMCID: PMC5784943
- DOI: 10.1371/journal.pone.0191617
Inhibition of EV71 by curcumin in intestinal epithelial cells
Abstract
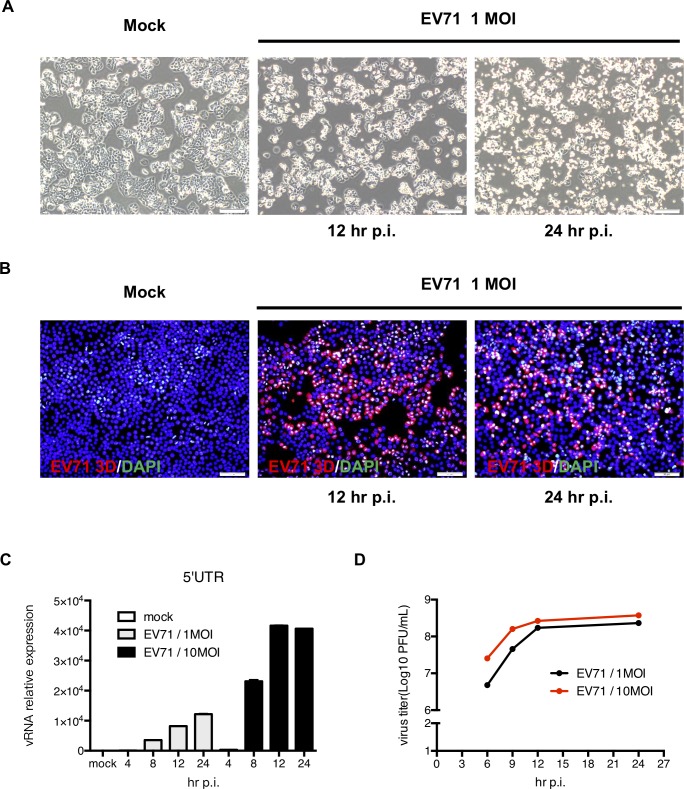
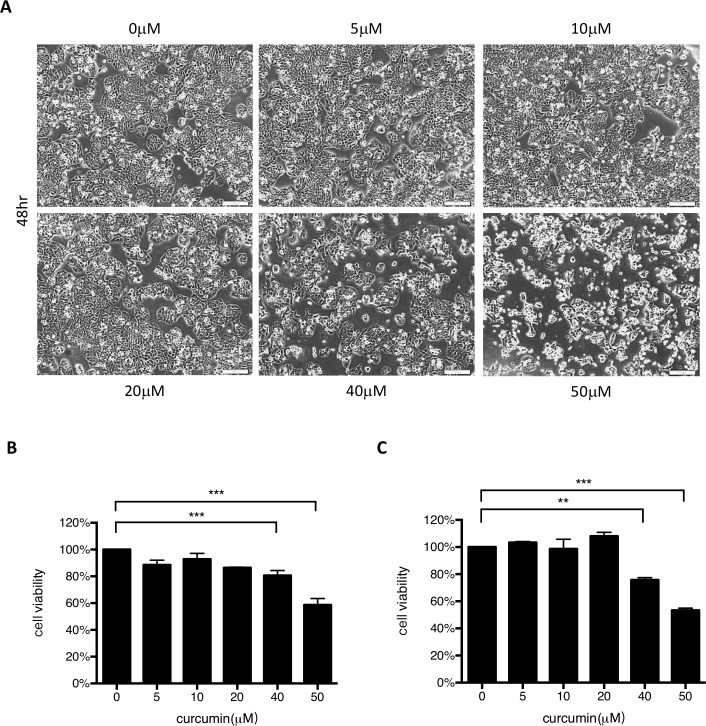
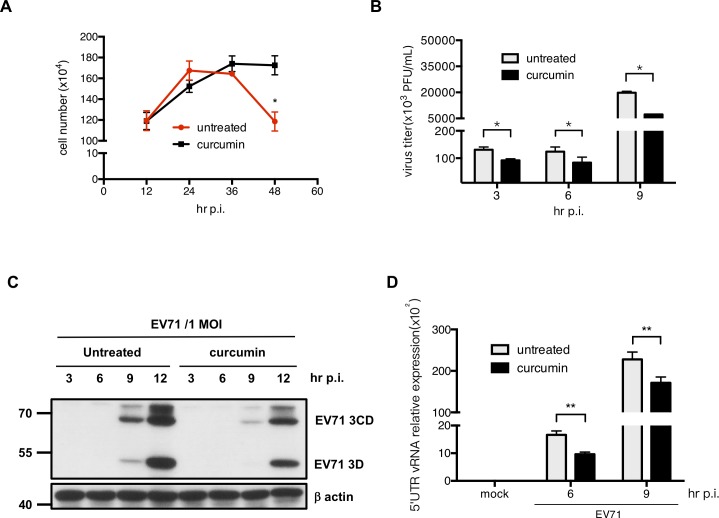
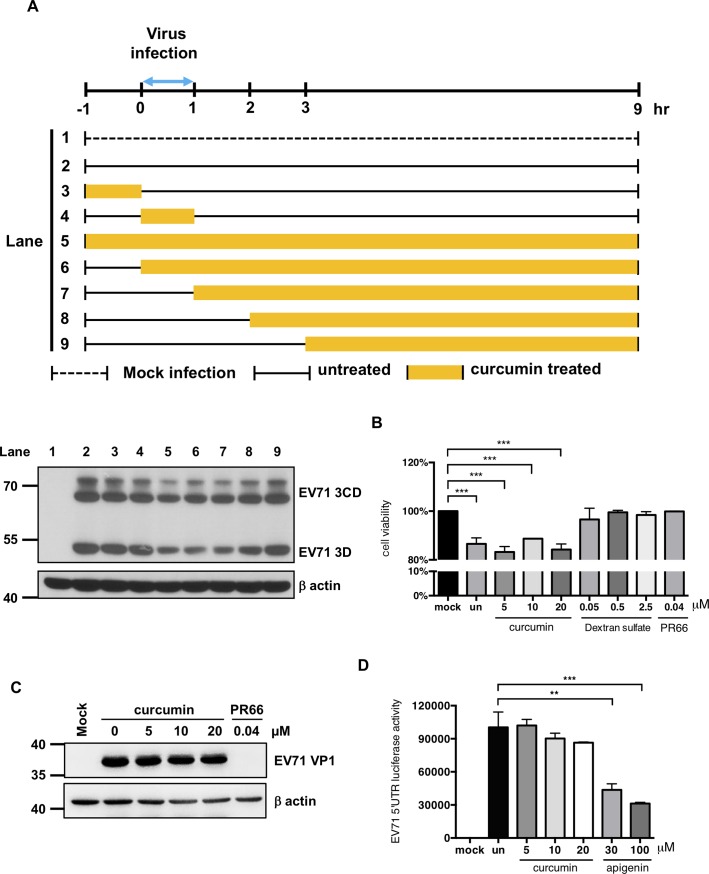
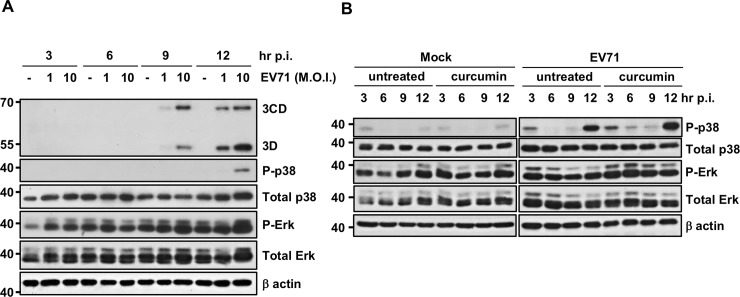
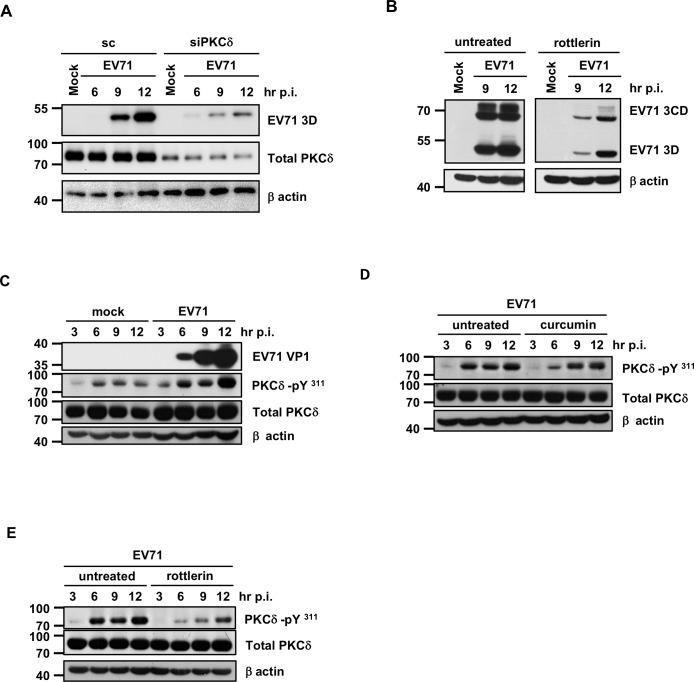
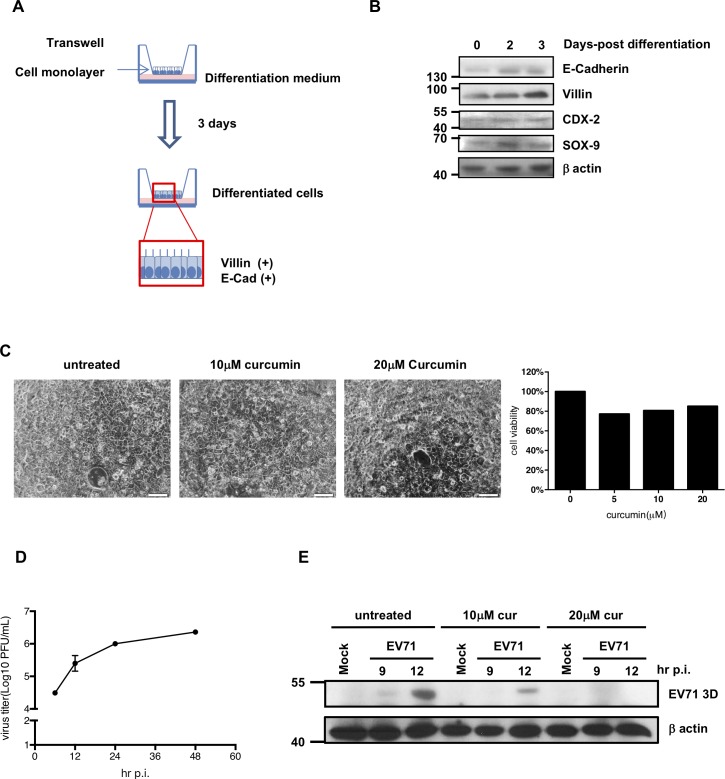
EV71 is a positive-sense single-stranded RNA virus that belongs to the Picornaviridae family. EV71 infection may cause various symptoms ranging from hand-foot-and-mouth disease to neurological pathological conditions such as aseptic meningitis, ataxia, and acute transverse myelitis. There is currently no effective treatment or vaccine available. Various compounds have been examined for their ability to restrict EV71 replication. However, most experiments have been performed in rhabdomyosarcoma or Vero cells. Since the gastrointestinal tract is the entry site for this pathogen, we anticipated that orally ingested agents may exert beneficial effects by decreasing virus replication in intestinal epithelial cells. In this study, curcumin (diferuloylmethane, C21H20O6), an active ingredient of turmeric (Curcuma longa Linn) with anti-cancer properties, was investigated for its anti-enterovirus activity. We demonstrate that curcumin treatment inhibits viral translation and increases host cell viability. Curcumin does not exert its anti-EV71 effects by modulating virus attachment or virus internal ribosome entry site (IRES) activity. Furthermore, curcumin-mediated regulation of mitogen-activated protein kinase (MAPK) signaling pathways is not involved. We found that protein kinase C delta (PKCδ) plays a role in virus translation in EV71-infected intestinal epithelial cells and that curcumin treatment decreases the phosphorylation of this enzyme. In addition, we show evidence that curcumin also limits viral translation in differentiated human intestinal epithelial cells. In summary, our data demonstrate the anti-EV71 properties of curcumin, suggesting that ingestion of this phytochemical may protect against enteroviral infections.
Conflict of interest statement
Figures
References
-
- Chan KP, Goh KT, Chong CY, Teo ES, Lau G, Ling AE. Epidemic hand, foot and mouth disease caused by human enterovirus 71, Singapore. Emerg Infect Dis. 2003; 9: 78–85. doi: 10.3201/eid1301.020112 - DOI - PMC - PubMed
-
- Shimizu H, Utama A, Onnimala N, Li C, Li-Bi Z, Yu-Jie M et al. Molecular epidemiology of enterovirus 71 infection in the Western Pacific Region. Pediatr Int. 2004; 46: 231–235. doi: 10.1046/j.1442-200x.2004.01868.x - DOI - PubMed
-
- Chan LG, Parashar UD, Lye MS, Ong FG, Zaki SR, Alexander JP et al. Deaths of children during an outbreak of hand, foot, and mouth disease in sarawak, malaysia: clinical and pathological characteristics of the disease. For the Outbreak Study Group. Clin Infect Dis. 2000; 31: 678–683. doi: 10.1086/314032 - DOI - PubMed
-
- Galasiti Kankanamalage AC, Kim Y, Weerawarna PM, Uy RA, Damalanka VC, Mandadapu SR et al. Structure-guided design and optimization of dipeptidyl inhibitors of norovirus 3CL protease. Structure-activity relationships and biochemical, X-ray crystallographic, cell-based, and in vivo studies. J Med Chem. 2015; 58: 3144–3155. doi: 10.1021/jm5019934 - DOI - PMC - PubMed
-
- Yang Y, Xiu J, Liu J, Zhang L, Li X, Xu Y et al. Chebulagic acid, a hydrolyzable tannin, exhibited antiviral activity in vitro and in vivo against human enterovirus 71. Int J Mol Sci. 2013; 14:9618–9627. doi: 10.3390/ijms14059618 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
