

Lipid droplet formation in Mycobacterium tuberculosis infected macrophages requires IFN-γ/HIF-1α signaling and supports host defense
- PMID: 29370315
- PMCID: PMC5800697
- DOI: 10.1371/journal.ppat.1006874
Lipid droplet formation in Mycobacterium tuberculosis infected macrophages requires IFN-γ/HIF-1α signaling and supports host defense
Abstract
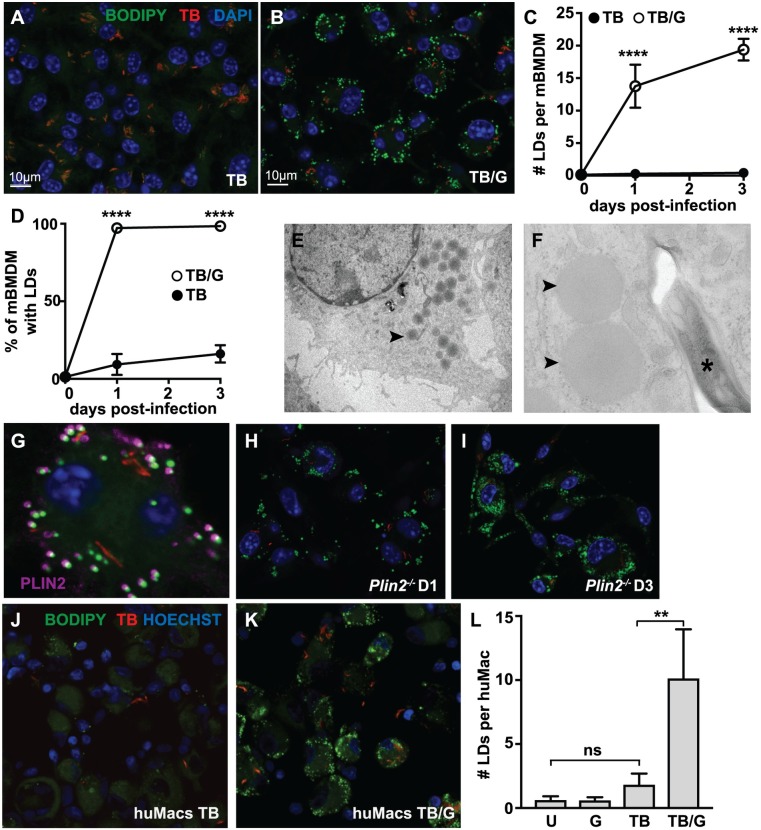
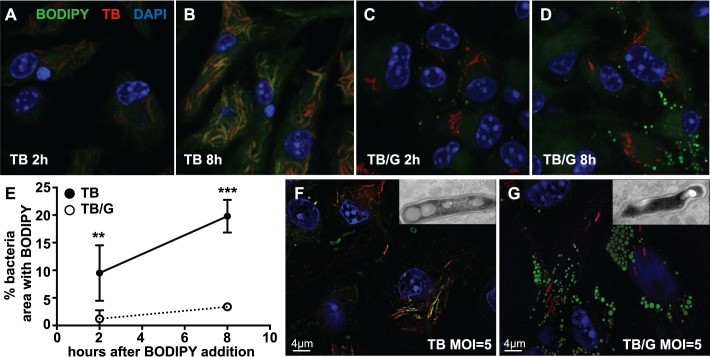
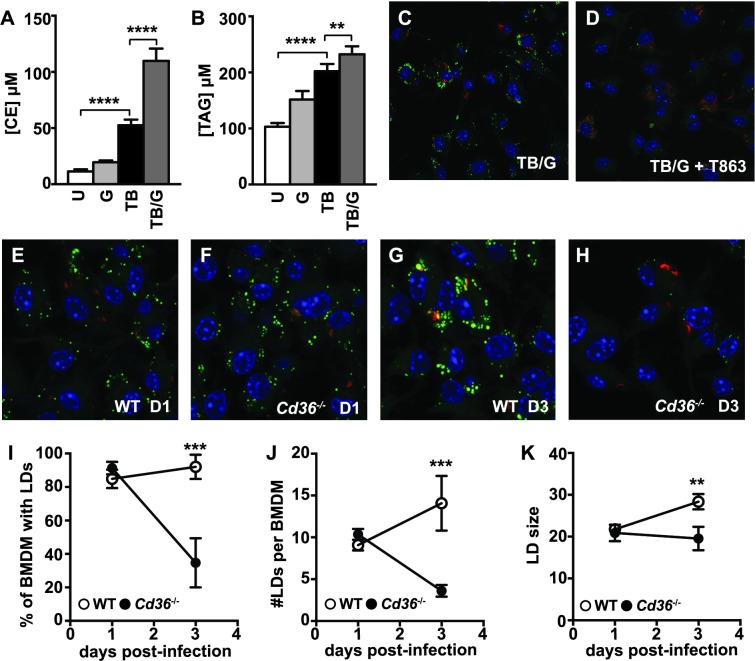
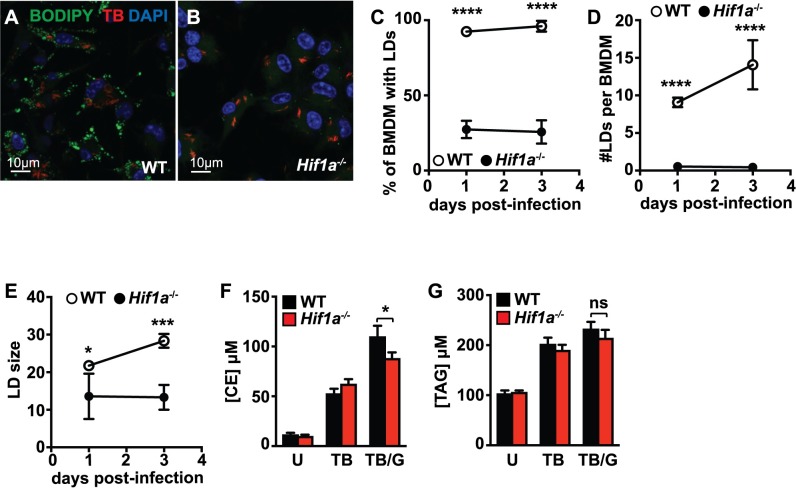
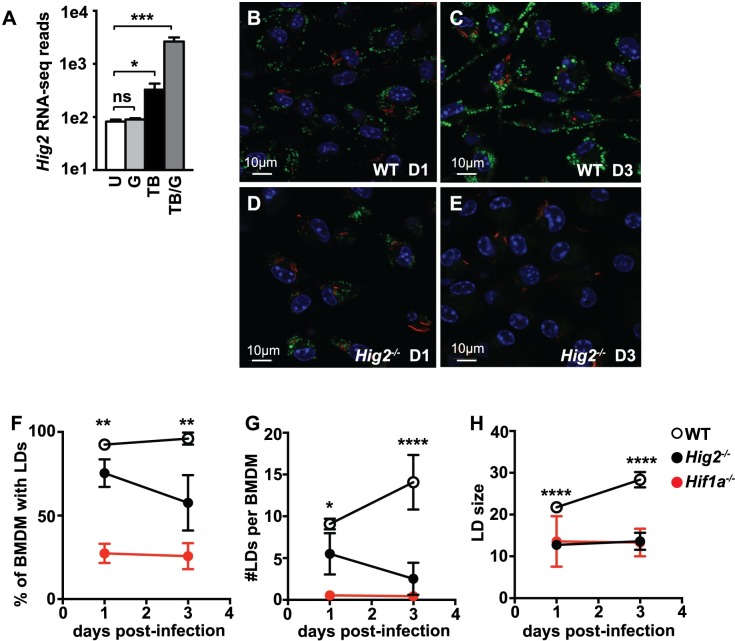
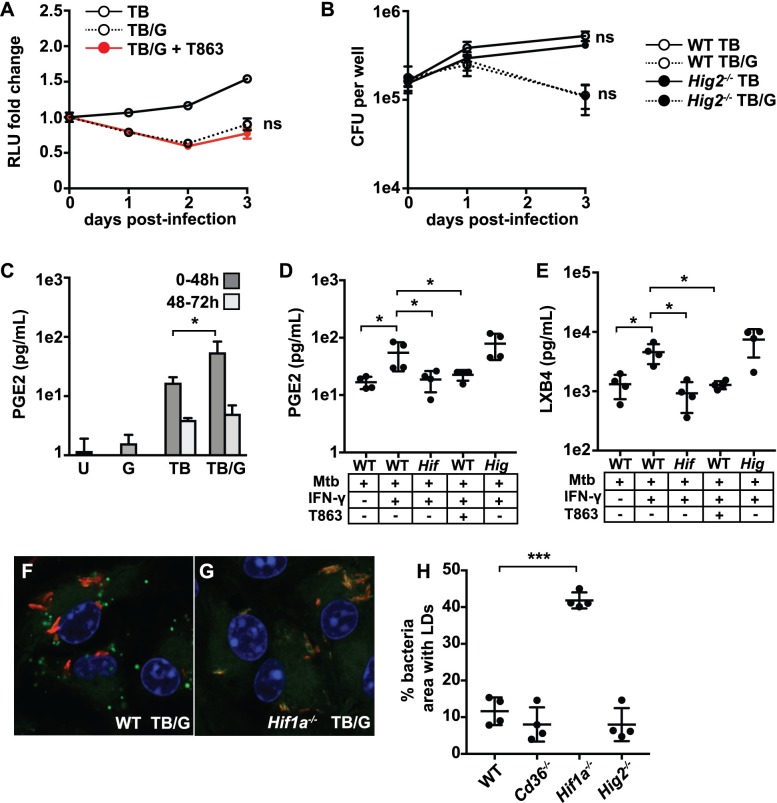
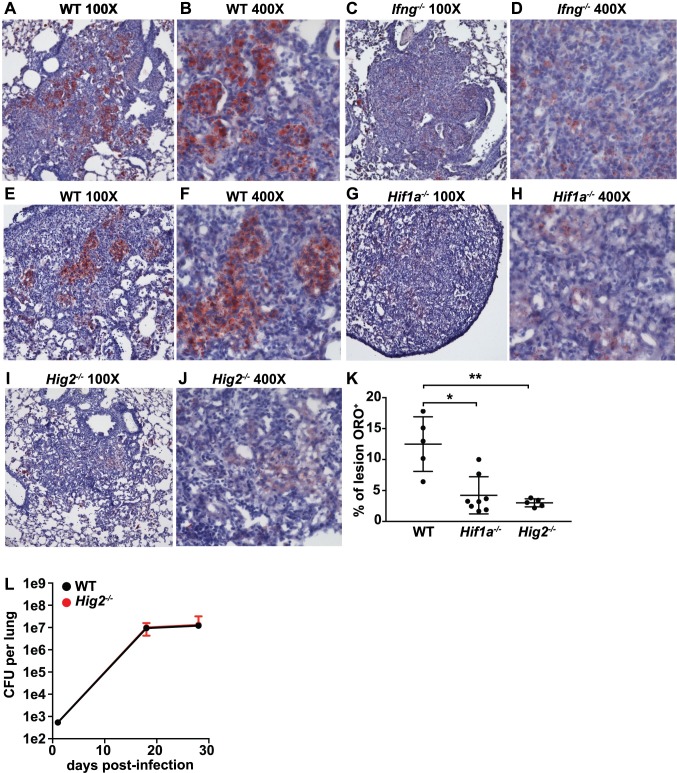
Lipid droplet (LD) formation occurs during infection of macrophages with numerous intracellular pathogens, including Mycobacterium tuberculosis. It is believed that M. tuberculosis and other bacteria specifically provoke LD formation as a pathogenic strategy in order to create a depot of host lipids for use as a carbon source to fuel intracellular growth. Here we show that LD formation is not a bacterially driven process during M. tuberculosis infection, but rather occurs as a result of immune activation of macrophages as part of a host defense mechanism. We show that an IFN-γ driven, HIF-1α dependent signaling pathway, previously implicated in host defense, redistributes macrophage lipids into LDs. Furthermore, we show that M. tuberculosis is able to acquire host lipids in the absence of LDs, but not in the presence of IFN-γ induced LDs. This result uncouples macrophage LD formation from bacterial acquisition of host lipids. In addition, we show that IFN-γ driven LD formation supports the production of host protective eicosanoids including PGE2 and LXB4. Finally, we demonstrate that HIF-1α and its target gene Hig2 are required for the majority of LD formation in the lungs of mice infected with M. tuberculosis, thus demonstrating that immune activation provides the primary stimulus for LD formation in vivo. Taken together our data demonstrate that macrophage LD formation is a host-driven component of the adaptive immune response to M. tuberculosis, and suggest that macrophage LDs are not an important source of nutrients for M. tuberculosis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
HIF-1α Is an Essential Mediator of IFN-γ-Dependent Immunity to Mycobacterium tuberculosis.J Immunol. 2016 Aug 15;197(4):1287-97. doi: 10.4049/jimmunol.1600266. Epub 2016 Jul 18. J Immunol. 2016. PMID: 27430718 Free PMC article.
-
Nitric Oxide Modulates Macrophage Responses to Mycobacterium tuberculosis Infection through Activation of HIF-1α and Repression of NF-κB.J Immunol. 2017 Sep 1;199(5):1805-1816. doi: 10.4049/jimmunol.1700515. Epub 2017 Jul 28. J Immunol. 2017. PMID: 28754681 Free PMC article.
-
IFN-γ-independent control of M. tuberculosis requires CD4 T cell-derived GM-CSF and activation of HIF-1α.PLoS Pathog. 2022 Jul 25;18(7):e1010721. doi: 10.1371/journal.ppat.1010721. eCollection 2022 Jul. PLoS Pathog. 2022. PMID: 35877763 Free PMC article.
-
Macrophage defense mechanisms against intracellular bacteria.Immunol Rev. 2015 Mar;264(1):182-203. doi: 10.1111/imr.12266. Immunol Rev. 2015. PMID: 25703560 Free PMC article. Review.
-
[Protective immunity against Mycobacterium tuberculosis].Kekkaku. 2006 Nov;81(11):687-91. Kekkaku. 2006. PMID: 17154048 Review. Japanese.
Cited by
-
Immunometabolism during Mycobacterium tuberculosis Infection.Trends Microbiol. 2020 Oct;28(10):832-850. doi: 10.1016/j.tim.2020.04.010. Epub 2020 May 11. Trends Microbiol. 2020. PMID: 32409147 Free PMC article. Review.
-
Mycobacterium tuberculosis and myeloid-derived suppressor cells: Insights into caveolin rich lipid rafts.EBioMedicine. 2020 Mar;53:102670. doi: 10.1016/j.ebiom.2020.102670. Epub 2020 Feb 26. EBioMedicine. 2020. PMID: 32113158 Free PMC article. Review.
-
Mycobacterium tuberculosis infection of host cells in space and time.FEMS Microbiol Rev. 2019 Jul 1;43(4):341-361. doi: 10.1093/femsre/fuz006. FEMS Microbiol Rev. 2019. PMID: 30916769 Free PMC article. Review.
-
Tribbles1 is host protective during in vivo mycobacterial infection.Elife. 2024 Jun 18;13:e95980. doi: 10.7554/eLife.95980. Elife. 2024. PMID: 38896446 Free PMC article.
-
Lipid droplets in pathogen infection and host immunity.Acta Pharmacol Sin. 2024 Mar;45(3):449-464. doi: 10.1038/s41401-023-01189-1. Epub 2023 Nov 22. Acta Pharmacol Sin. 2024. PMID: 37993536 Free PMC article. Review.
References
-
- Floyd K. Global Tuberculosis Report 2016. Geneva: World Health Organization; 2016. December pp. 1–214.
-
- Getahun H, Matteelli A, Chaisson RE, Raviglione M. Latent Mycobacterium tuberculosis infection. The New England journal of medicine. 2015;372: 2127–2135. doi: 10.1056/NEJMra1405427 - DOI - PubMed
-
- Peyron P, Vaubourgeix J, Poquet Y, Levillain F, Botanch C, Bardou F, et al. Foamy macrophages from tuberculous patients' granulomas constitute a nutrient-rich reservoir for M. tuberculosis persistence. PLOS Pathogens. 2008;4: e1000204 doi: 10.1371/journal.ppat.1000204 - DOI - PMC - PubMed
-
- Russell DG, Cardona P-J, Kim M-J, Allain S, Altare F. Foamy macrophages and the progression of the human tuberculosis granuloma. Nat Immunol. 2009;10: 943–948. doi: 10.1038/ni.1781 - DOI - PMC - PubMed
-
- Kim M-J, Wainwright HC, Locketz M, Bekker L-G, Walther GB, Dittrich C, et al. Caseation of human tuberculosis granulomas correlates with elevated host lipid metabolism. EMBO Mol Med. 2010;2: 258–274. doi: 10.1002/emmm.201000079 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
