

Immunization with Transgenic Rodent Malaria Parasites Expressing Pfs25 Induces Potent Transmission-Blocking Activity
- PMID: 29371619
- PMCID: PMC5785477
- DOI: 10.1038/s41598-017-18831-8
Immunization with Transgenic Rodent Malaria Parasites Expressing Pfs25 Induces Potent Transmission-Blocking Activity
Abstract
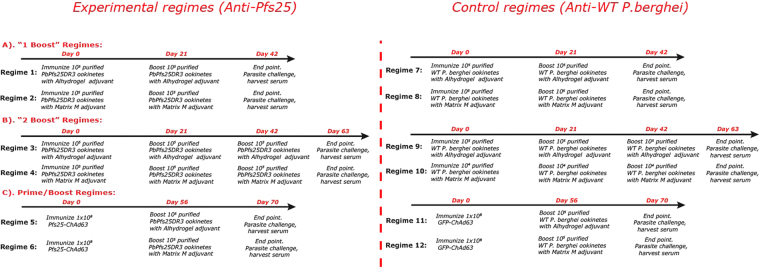
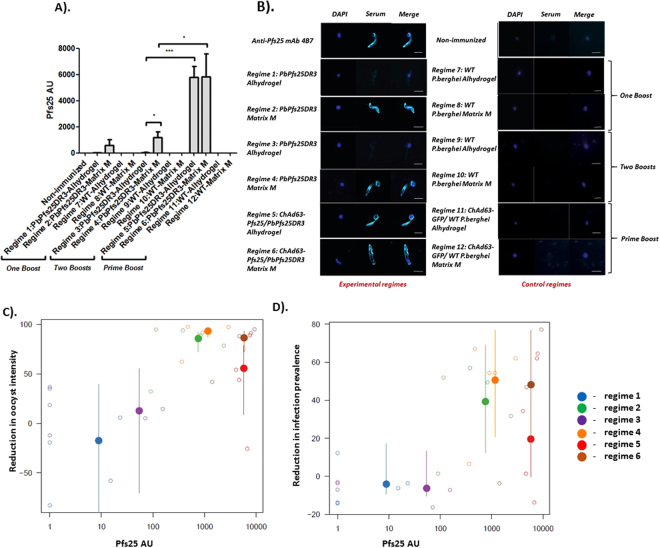
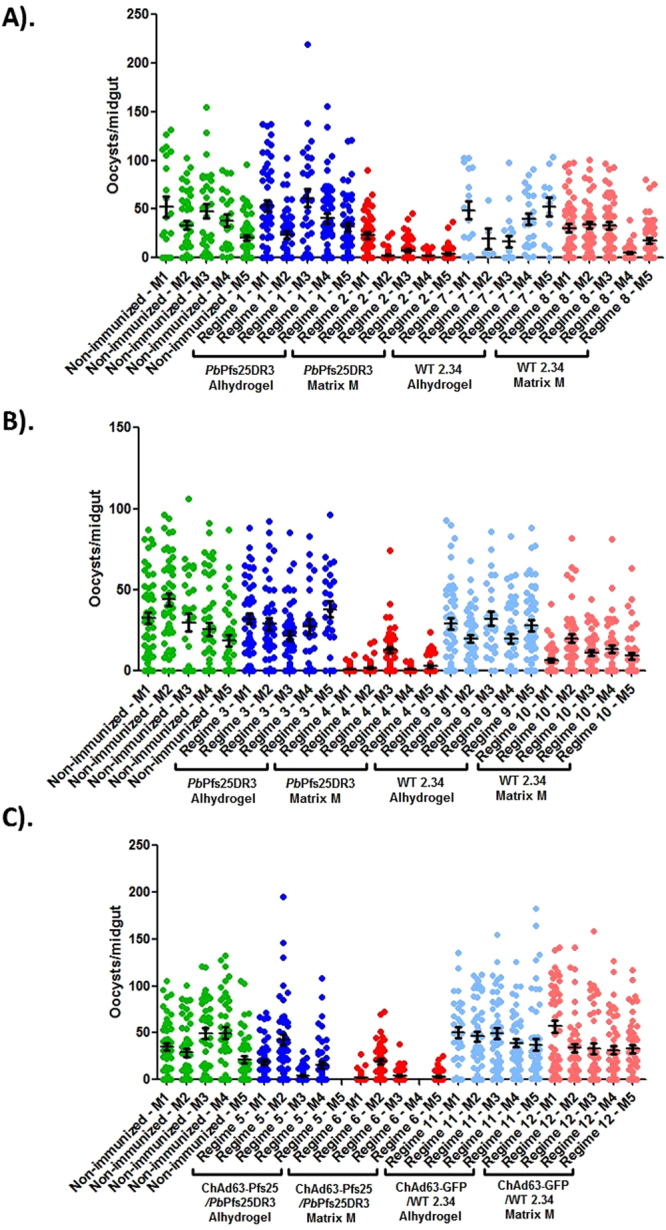
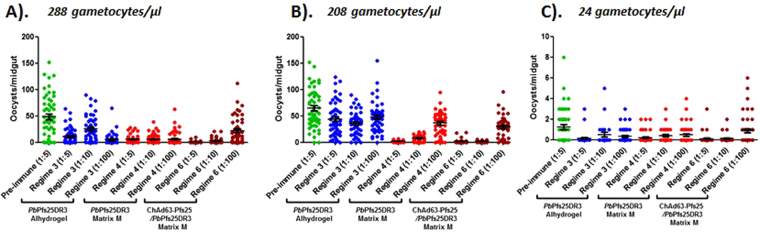
An anti-malarial transmission blocking vaccine (TBV) would be an important tool for disease control or elimination, though current candidates have failed to induce high efficacy in clinical studies. The ookinete surface protein P25 is a primary target for TBV development, but heterologous expression of P25 with appropriate conformation is problematic and a pre-requisite for achieving functional titers. A potential alternative to recombinant/sub-unit vaccine is immunization with a non-pathogenic, whole-parasite vaccine. This study examines the ability of a purified transgenic rodent-malaria parasite (PbPfs25DR3), expressing Plasmodium falciparum P25 in native conformation on the P. berghei ookinete surface, to act as a TBV. Vaccination with purified PbPfs25DR3 ookinetes produces a potent anti-Pfs25 response and high transmission-blocking efficacy in the laboratory, findings that are then translated to experimentation on natural field isolates of P. falciparum from infected individuals in Burkina Faso. Efficacy is demonstrated in the lab and the field (up to 93.3%/97.1% reductions in transmission intensity respectively), with both a homologous strategy with one and two boosts, and as part of a prime-boost regime, providing support for the future development of a whole-parasite TBV.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- WHO World Malaria report. World Health Organisation, http://www.who.int/malaria/publications/world_malaria_report/en/ (2017).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
