

Whole-Body Single-Cell Sequencing Reveals Transcriptional Domains in the Annelid Larval Body
- PMID: 29373712
- PMCID: PMC5913682
- DOI: 10.1093/molbev/msx336
Whole-Body Single-Cell Sequencing Reveals Transcriptional Domains in the Annelid Larval Body
Abstract
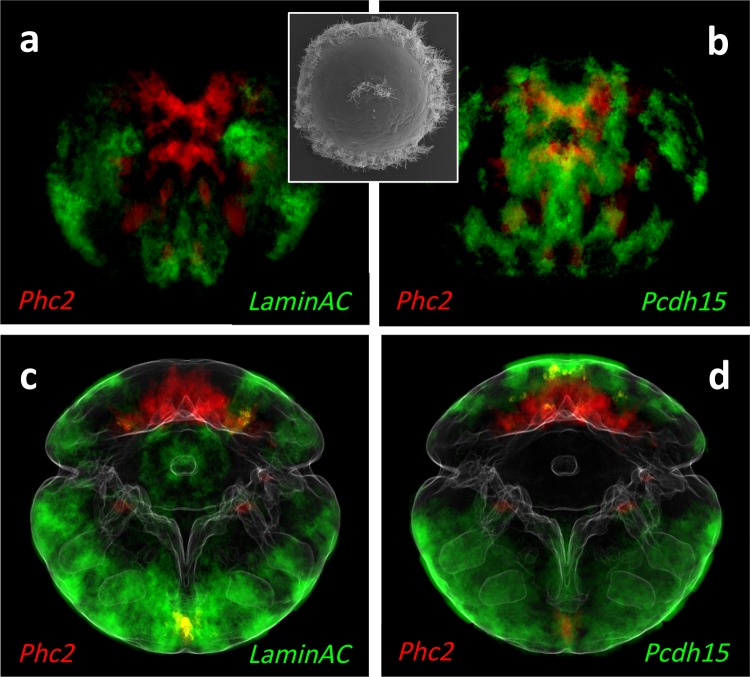
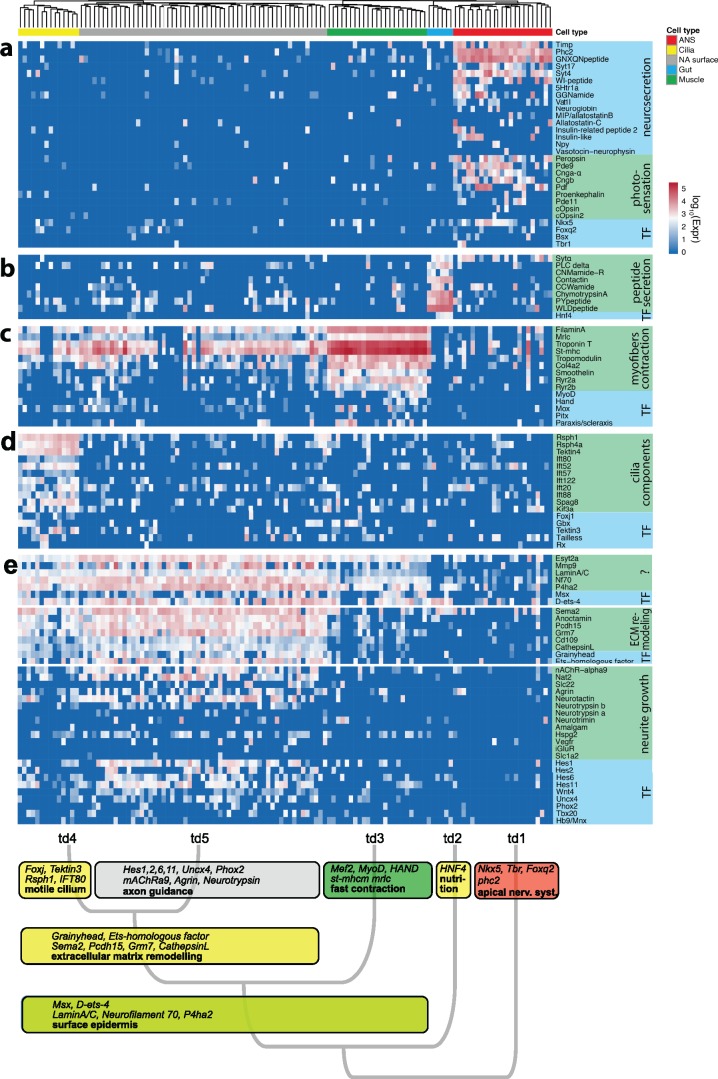
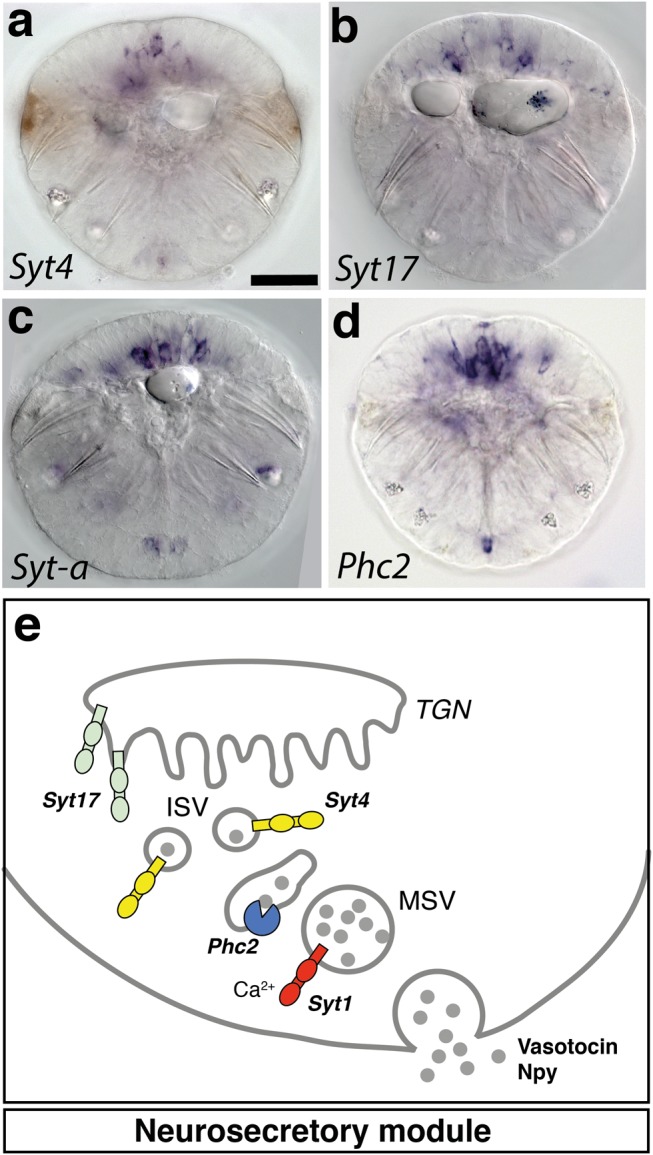
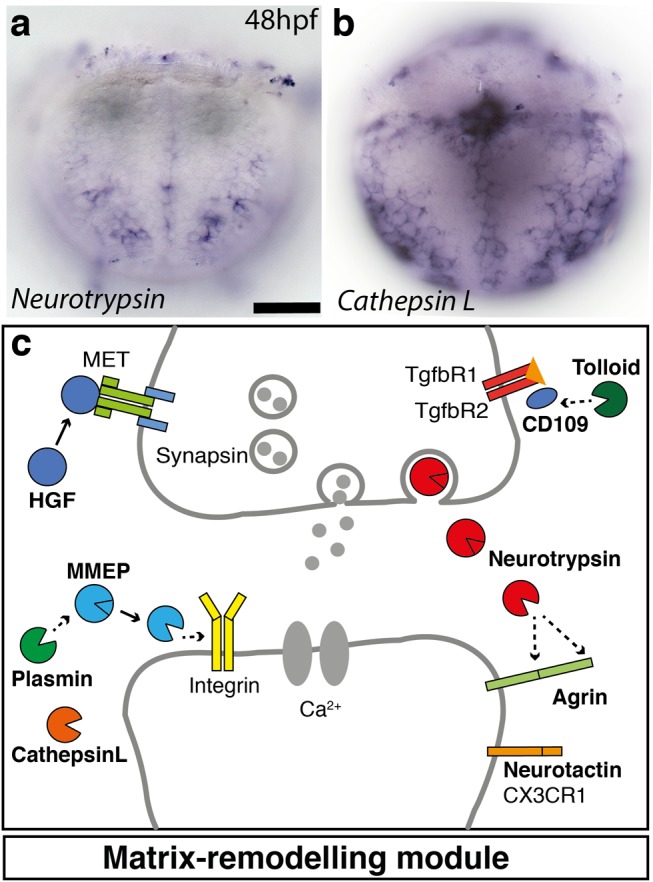
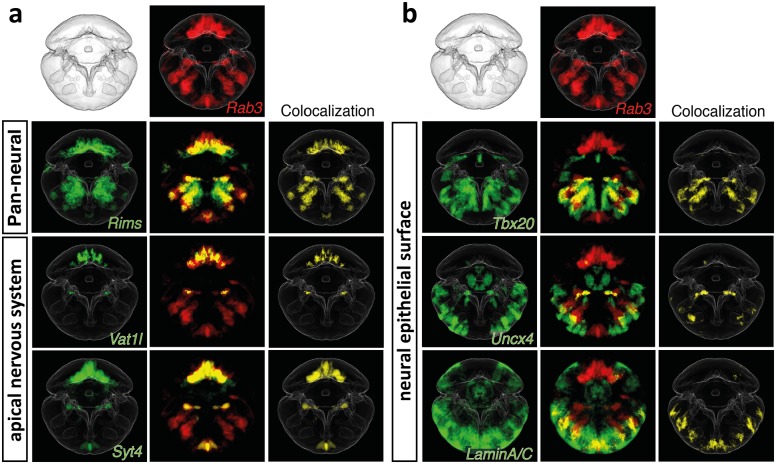
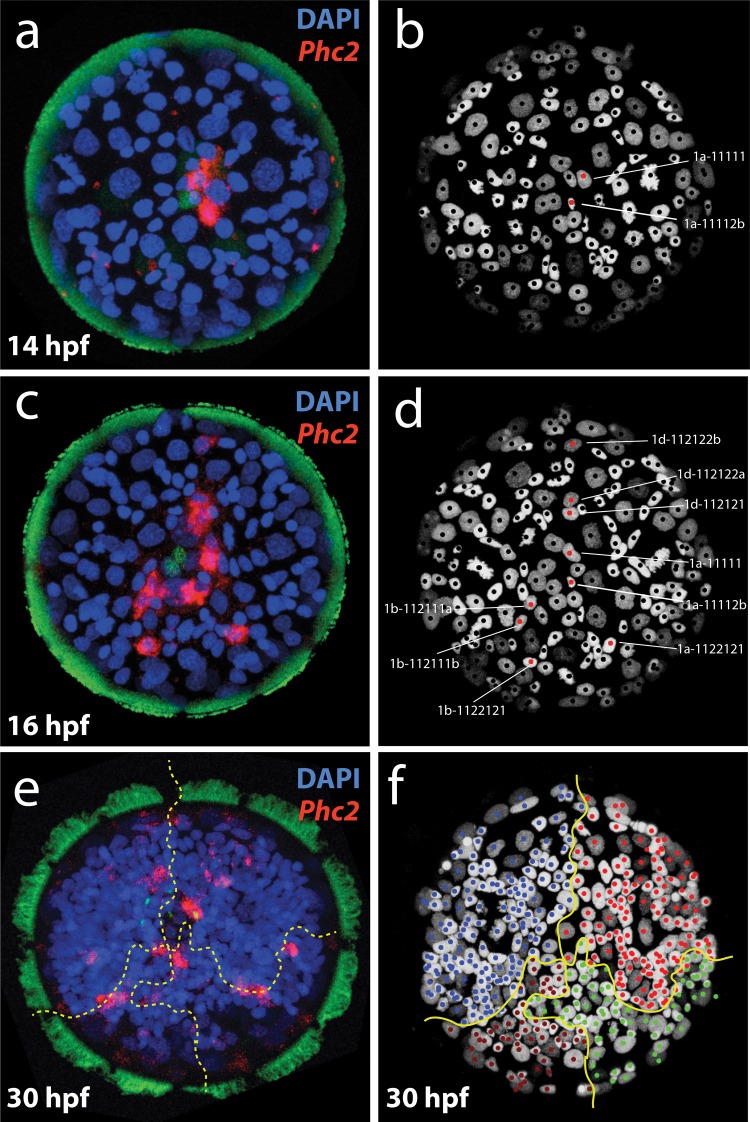
Animal bodies comprise diverse arrays of cells. To characterize cellular identities across an entire body, we have compared the transcriptomes of single cells randomly picked from dissociated whole larvae of the marine annelid Platynereis dumerilii. We identify five transcriptionally distinct groups of differentiated cells, each expressing a unique set of transcription factors and effector genes that implement cellular phenotypes. Spatial mapping of cells into a cellular expression atlas, and wholemount in situ hybridization of group-specific genes reveals spatially coherent transcriptional domains in the larval body, comprising, for example, apical sensory-neurosecretory cells versus neural/epidermal surface cells. These domains represent new, basic subdivisions of the annelid body based entirely on differential gene expression, and are composed of multiple, transcriptionally similar cell types. They do not represent clonal domains, as revealed by developmental lineage analysis. We propose that the transcriptional domains that subdivide the annelid larval body represent families of related cell types that have arisen by evolutionary diversification. Their possible evolutionary conservation makes them a promising tool for evo-devo research.
Figures
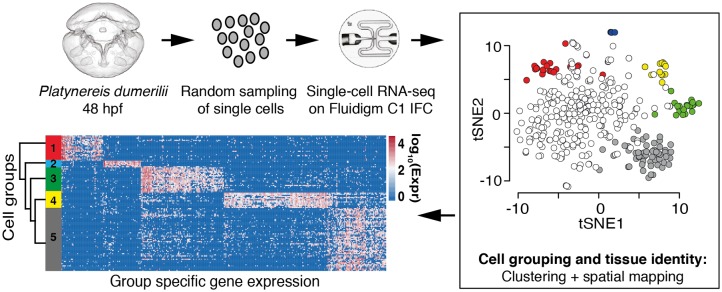
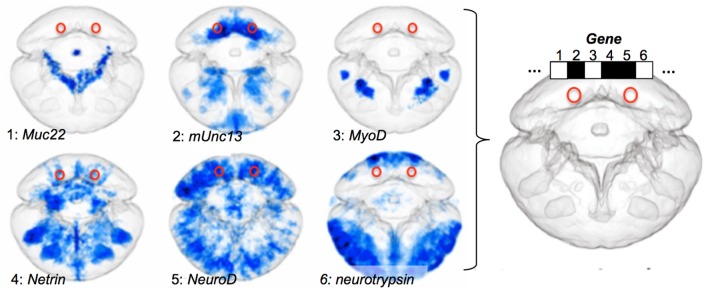
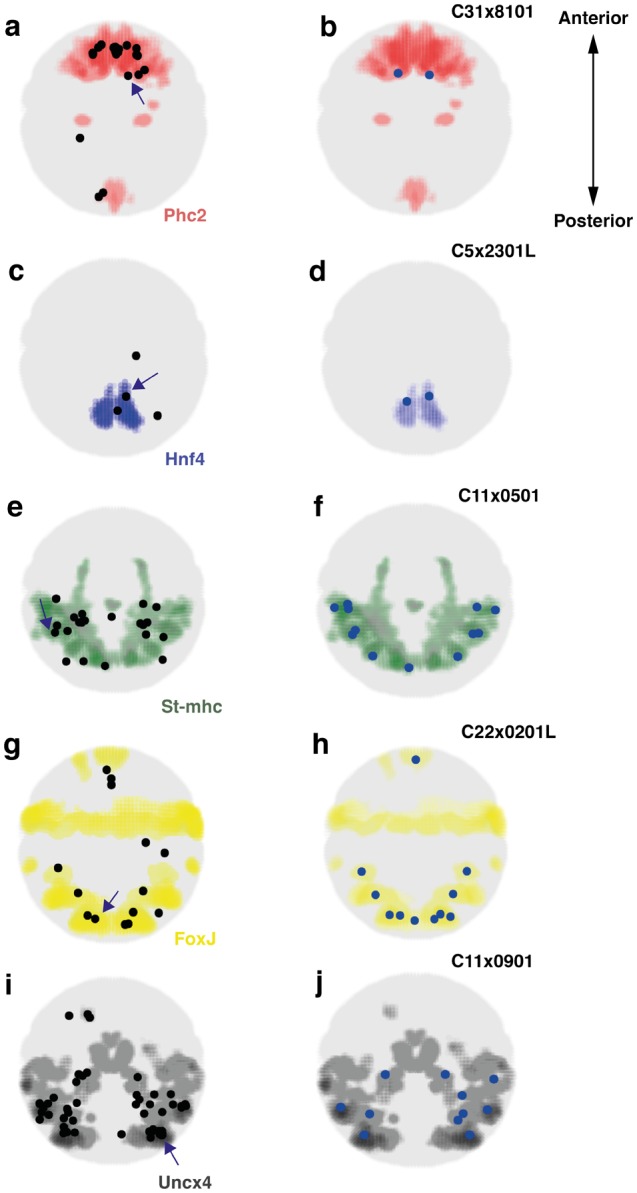
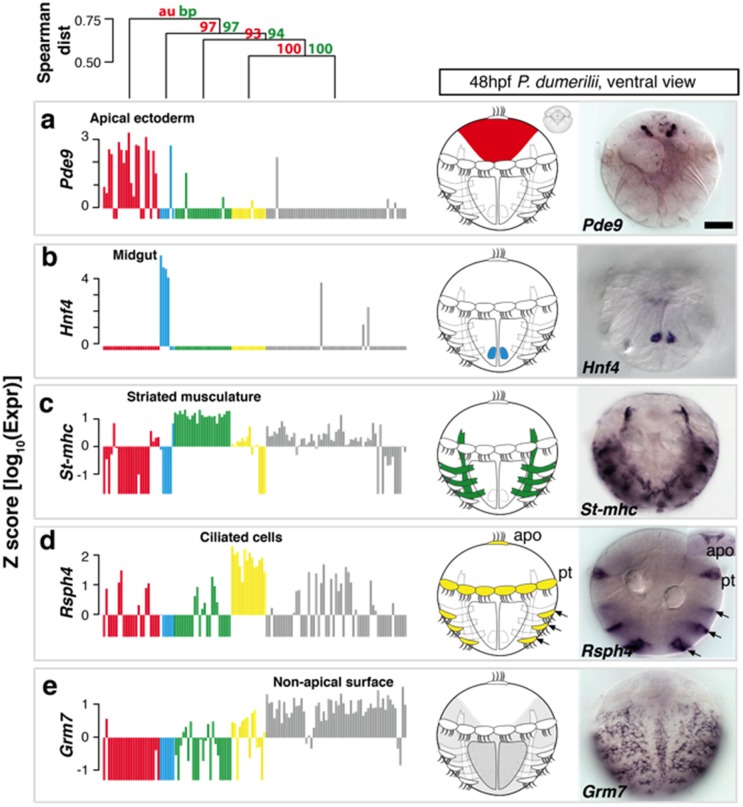
References
-
- Achim K, Pettit J-B, Saraiva LR, Gavriouchkina D, Larsson T, Arendt D, Marioni JC.. 2015. High-throughput spatial mapping of single-cell RNA-seq data to tissue of origin. Nat Biotechnol. 335:503–509. - PubMed
-
- Ackermann C, Dorresteijn A, Fischer A.. 2005. Clonal domains in postlarval Platynereis dumerilii (Annelida: Polychaeta). J Morphol. 2663:258–280. - PubMed
-
- Anders S, Pyl PT, Huber W.. 2015. HTSeq–a Python framework to work with high-throughput sequencing data. Bioinformatics 312:166–169.http://dx.doi.org/10.1093/bioinformatics/btu638 - DOI - PMC - PubMed
-
- Andrews S. 2010. FastQC: a quality control tool for high throughput sequence data. Available online at:http://www.bioinformatics.babraham.ac.uk/projects/fastqc/
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
