

CD22 Promotes B-1b Cell Responses to T Cell-Independent Type 2 Antigens
- PMID: 29374074
- PMCID: PMC5821584
- DOI: 10.4049/jimmunol.1701578
CD22 Promotes B-1b Cell Responses to T Cell-Independent Type 2 Antigens
Abstract
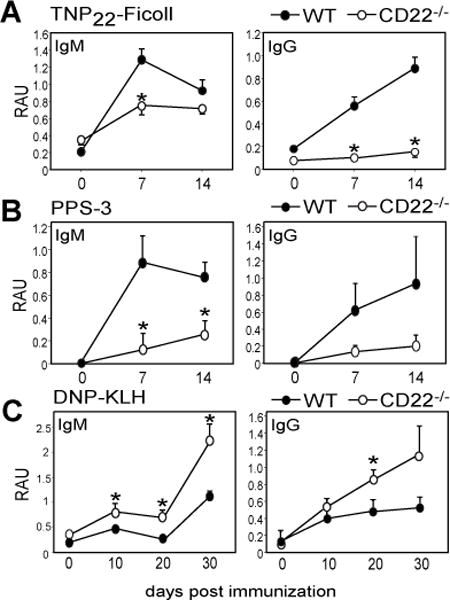
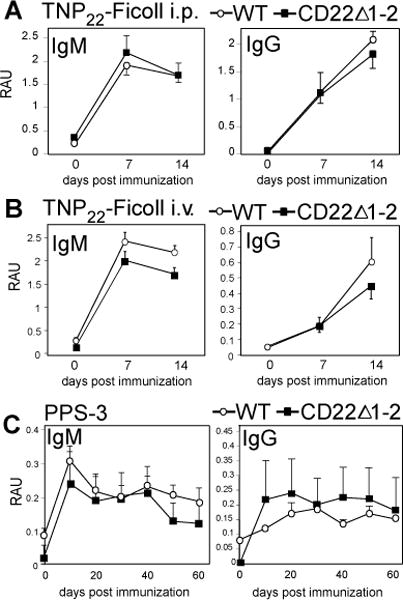
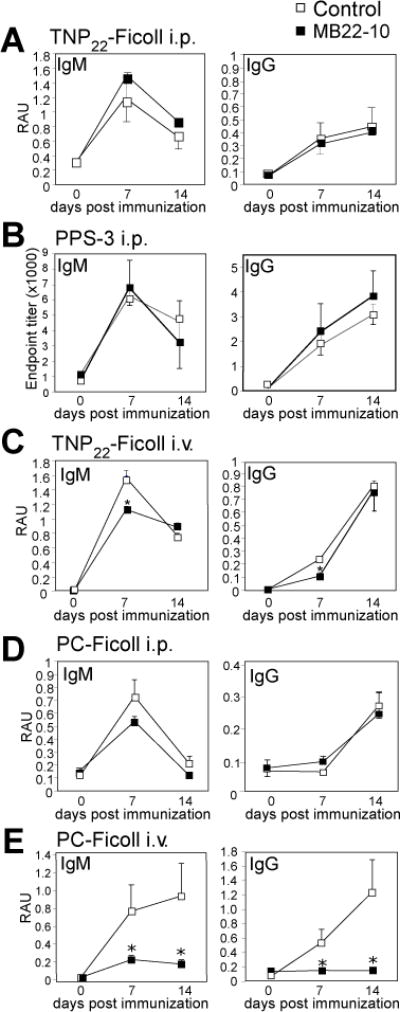
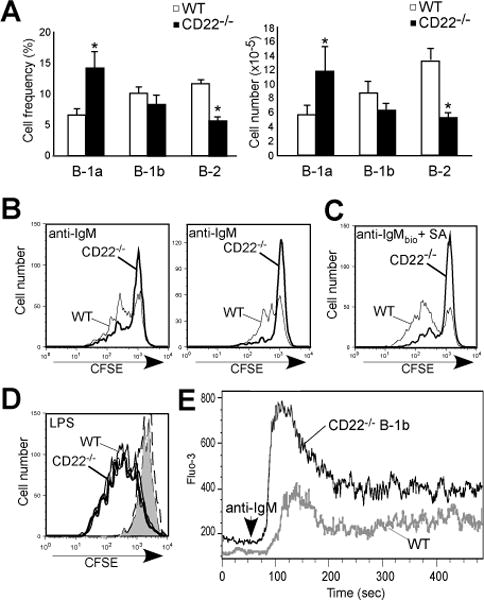
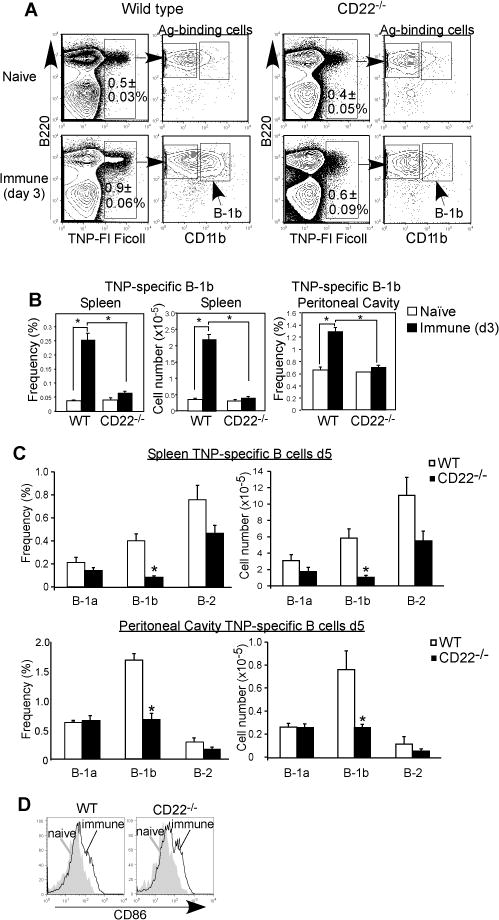
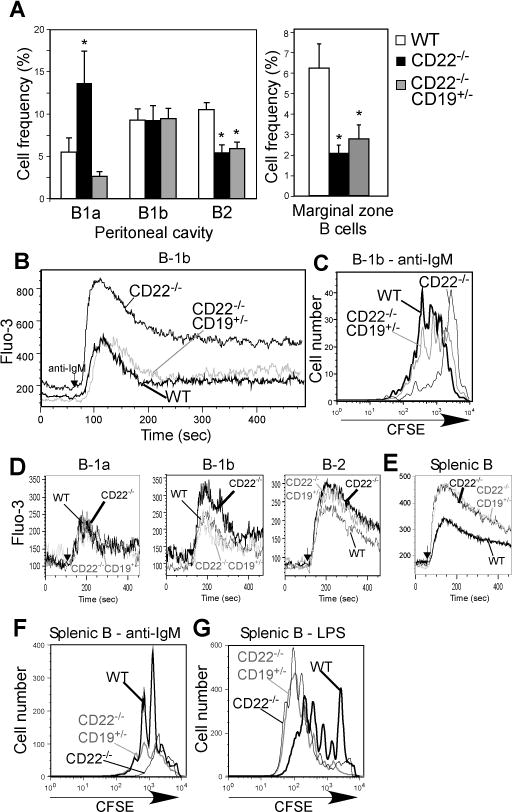
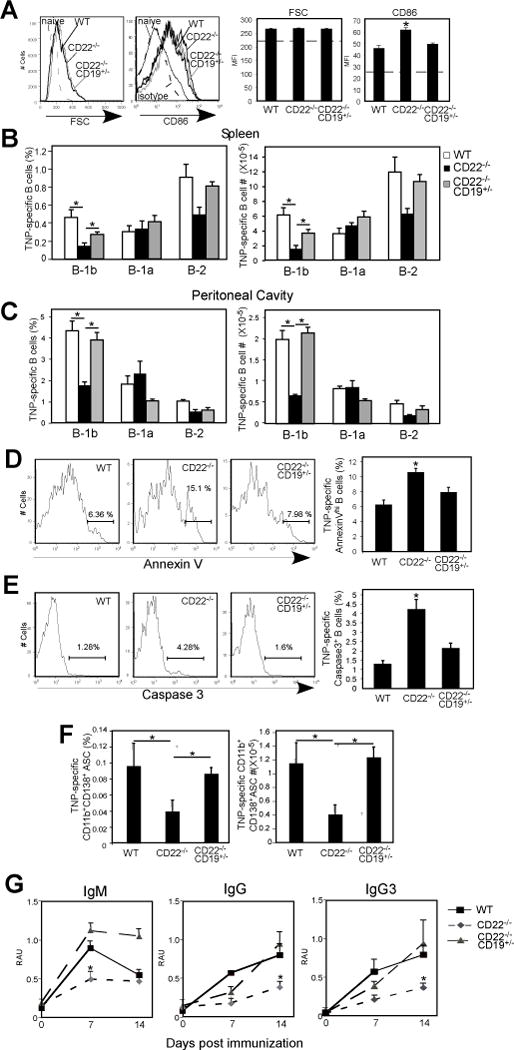
CD22 (Siglec-2) is a critical regulator of B cell activation and survival. CD22-/- mice generate significantly impaired Ab responses to T cell-independent type 2 (TI-2) Ags, including haptenated Ficoll and pneumococcal polysaccharides, Ags that elicit poor T cell help and activate BCR signaling via multivalent epitope crosslinking. This has been proposed to be due to impaired marginal zone (MZ) B cell development/maintenance in CD22-/- mice. However, mice expressing a mutant form of CD22 unable to bind sialic acid ligands generated normal TI-2 Ab responses, despite significantly reduced MZ B cells. Moreover, mice treated with CD22 ligand-binding blocking mAbs, which deplete MZ B cells, had little effect on TI-2 Ab responses. We therefore investigated the effects of CD22 deficiency on B-1b cells, an innate-like B cell population that plays a key role in TI-2 Ab responses. B-1b cells from CD22-/- mice had impaired BCR-induced proliferation and significantly increased intracellular Ca2+ concentration responses following BCR crosslinking. Ag-specific B-1b cell expansion and plasmablast differentiation following TI-2 Ag immunization was significantly impaired in CD22-/- mice, consistent with reduced TI-2 Ab responses. We generated CD22-/- mice with reduced CD19 levels (CD22-/-CD19+/-) to test the hypothesis that augmented B-1b cell BCR signaling in CD22-/- mice contributes to impaired TI-2 Ab responses. BCR-induced proliferation and intracellular Ca2+ concentration responses were normalized in CD22-/-CD19+/- B-1b cells. Consistent with this, TI-2 Ag-specific B-1b cell expansion, plasmablast differentiation, survival, and Ab responses were rescued in CD22-/-CD19+/- mice. Thus, CD22 plays a critical role in regulating TI-2 Ab responses through regulating B-1b cell signaling thresholds.
Copyright © 2018 by The American Association of Immunologists, Inc.
Figures
References
-
- Mehr S, Wood N. Streptococcus pneumoniae–a review of carriage, infection, serotype replacement and vaccination. Paediatric Resp Rev. 2012;13:258–64. - PubMed
-
- Vos Q, Lees A, Wu ZQ, Snapper CM, Mond JJ. B-cell activation by T-cell-independent type 2 antigens as an integral part of the humoral immune response to pathogenic microorganisms. Immunol Rev. 2000;176:154–170. - PubMed
-
- Gonzalez-Fernandez A, Faro J, Fernandez C. Immune responses to polysaccharides: lessons from humans and mice. Vaccine. 2008;26:292–300. - PubMed
-
- Weller S, Reynaud CA, Weill JC. Vaccination against encapsulated bacteria in humans: paradoxes. Trends Immunol. 2005;26:85–89. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
