

Grapevine VpPR10.1 functions in resistance to Plasmopara viticola through triggering a cell death-like defence response by interacting with VpVDAC3
- PMID: 29377445
- PMCID: PMC6041444
- DOI: 10.1111/pbi.12891
Grapevine VpPR10.1 functions in resistance to Plasmopara viticola through triggering a cell death-like defence response by interacting with VpVDAC3
Abstract
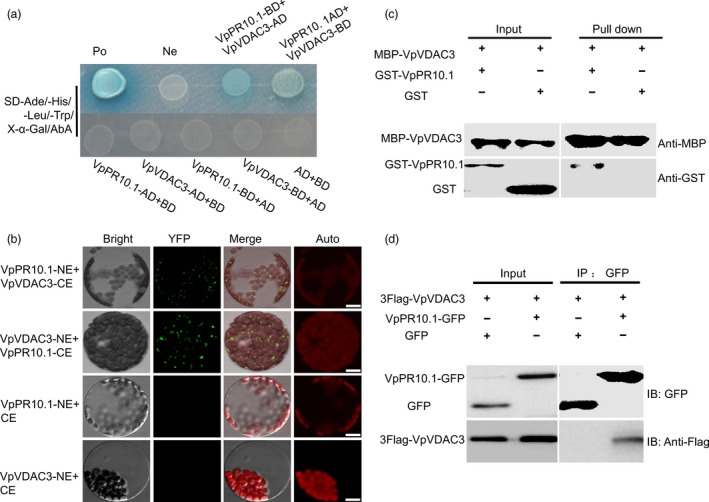
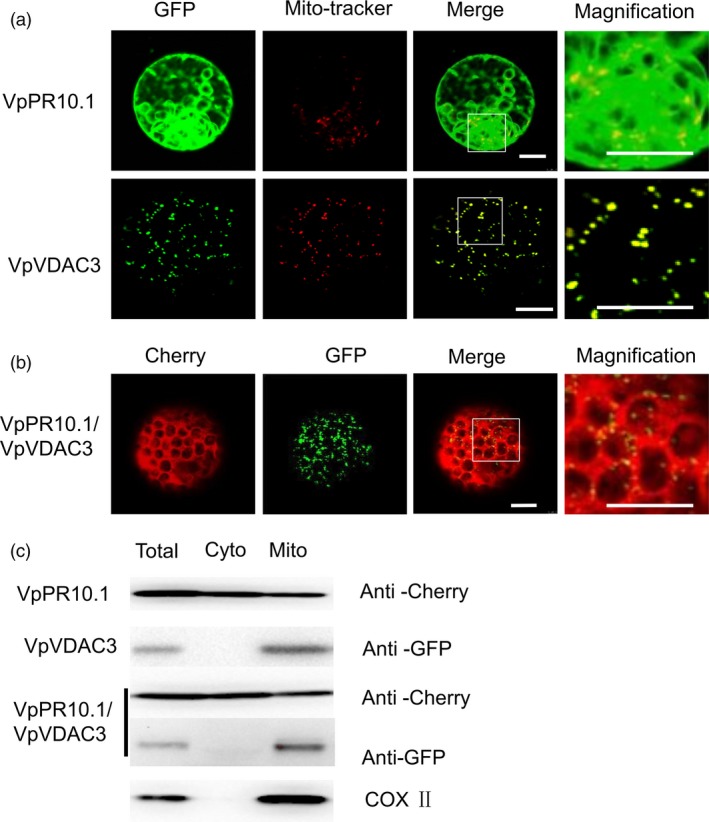
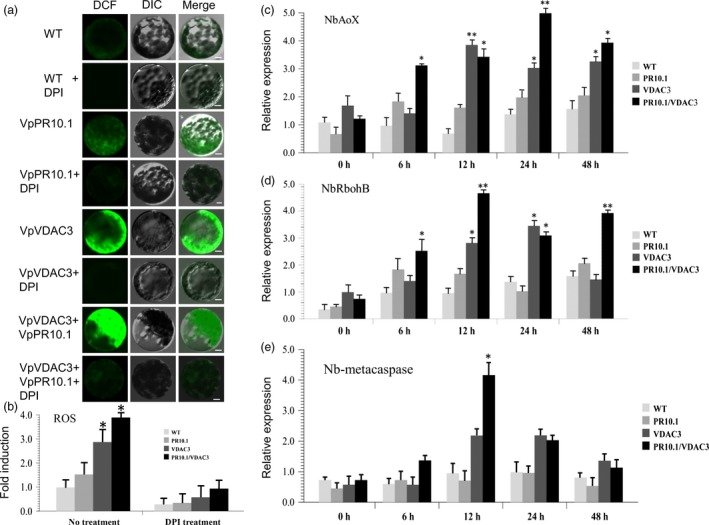
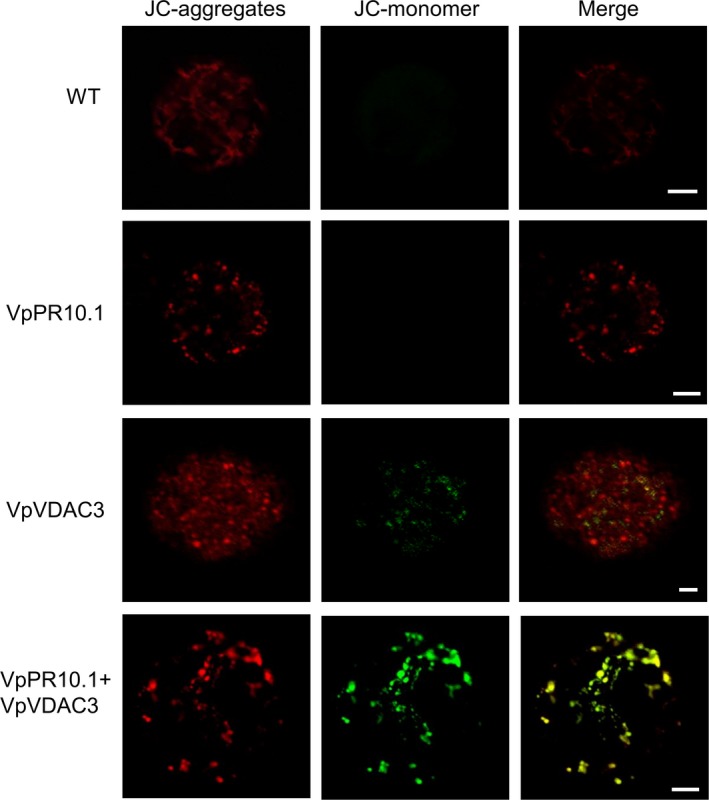
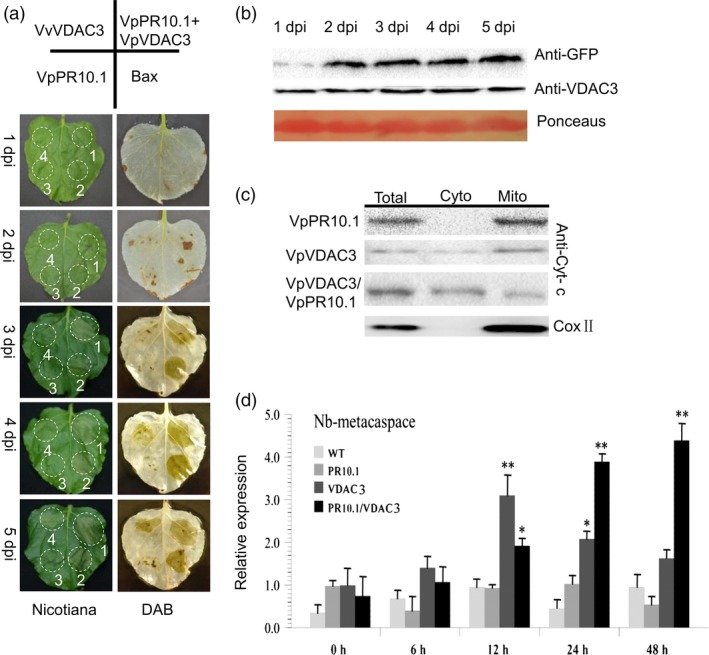
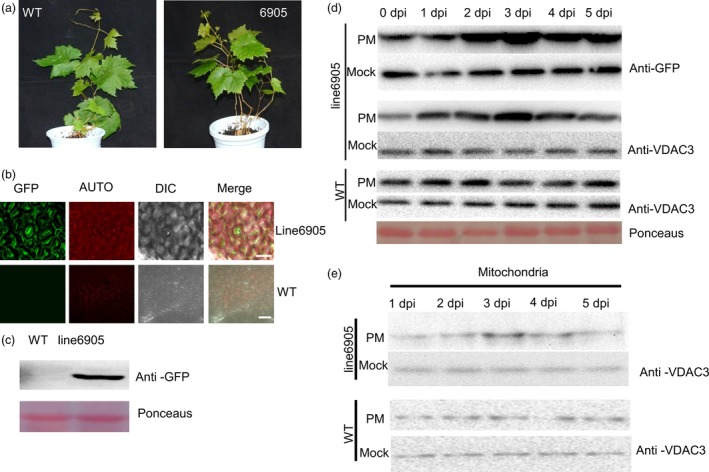
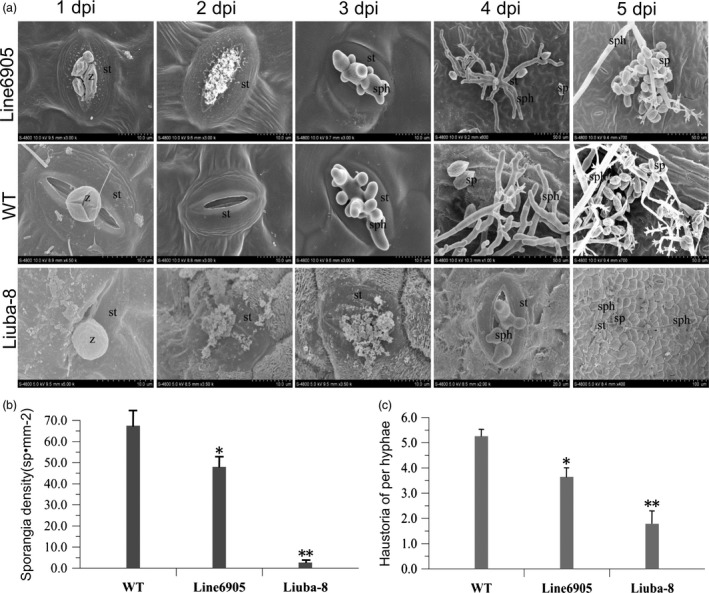
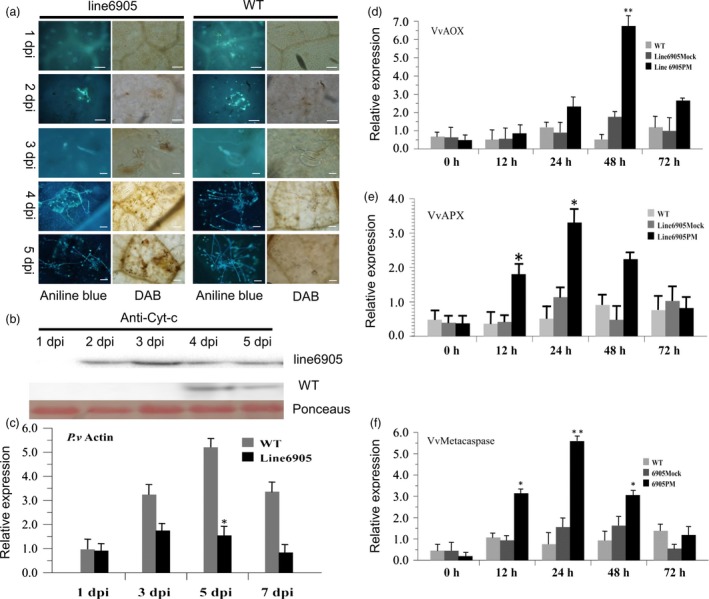
As one of the most serious diseases in grape, downy mildew caused by Plasmopara viticola is a worldwide grape disease. Much effort has been focused on improving susceptible grapevine resistance, and wild resistant grapevine species are important for germplasm improvement of commercial cultivars. Using yeast two-hybrid screen followed by a series of immunoprecipitation experiments, we identified voltage-dependent anion channel 3 (VDAC3) protein from Vitis piasezkii 'Liuba-8' as an interacting partner of VpPR10.1 cloned from Vitis pseudoreticulata 'Baihe-35-1', which is an important germplasm for its resistance to a range of pathogens. Co-expression of VpPR10.1/VpVDAC3 induced cell death in Nicotiana benthamiana, which accompanied by ROS accumulation. VpPR10.1 transgenic grapevine line showed resistance to P. viticola. We conclude that the VpPR10.1/VpVDAC3 complex is responsible for cell death-mediated defence response to P. viticola in grapevine.
Keywords: Plasmopara viticola; ROS; Vitis pseudoreticulata; PR10; grapevine; voltage-dependent anion channel.
© 2018 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Figures
References
-
- Al Bitar, F. , Roosens, N. , Smeyers, M. , Vauterin, M. , Van Boxtel, J. , Jacobs, M. and Homblé, F. (2003) Sequence analysis, transcriptional and posttranscriptional regulation of the rice vdac family. Biochim. Biophys. Acta. 1625, 43–51. - PubMed
-
- Benz, R. (1994) Permeation of hydrophilic solutes through mitochondrial outer membranes, review on mitochondrial porins. Biochem. Biophys. Acta. 1197, 167–196. - PubMed
-
- Bittel, P. and Robatzek, S. (2007) Microbe‐associated molecular patterns (MAMPs) probe plant immunity. Curr. Opin. Plant Biol. 10, 335–341. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
