

Improved insect-proofing: expressing double-stranded RNA in chloroplasts
- PMID: 29377554
- PMCID: PMC6055657
- DOI: 10.1002/ps.4870
Improved insect-proofing: expressing double-stranded RNA in chloroplasts
Abstract
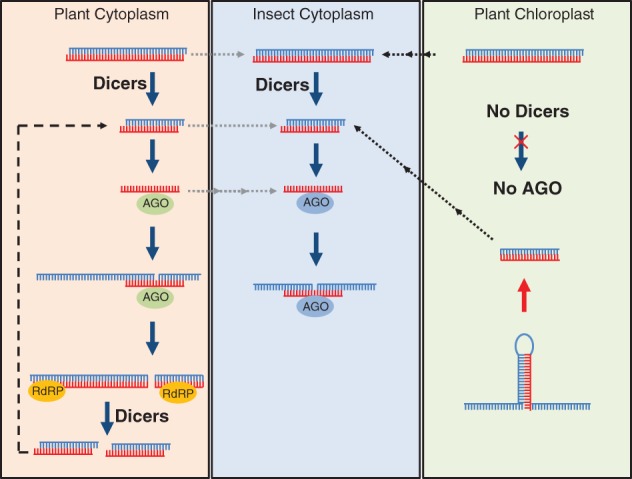
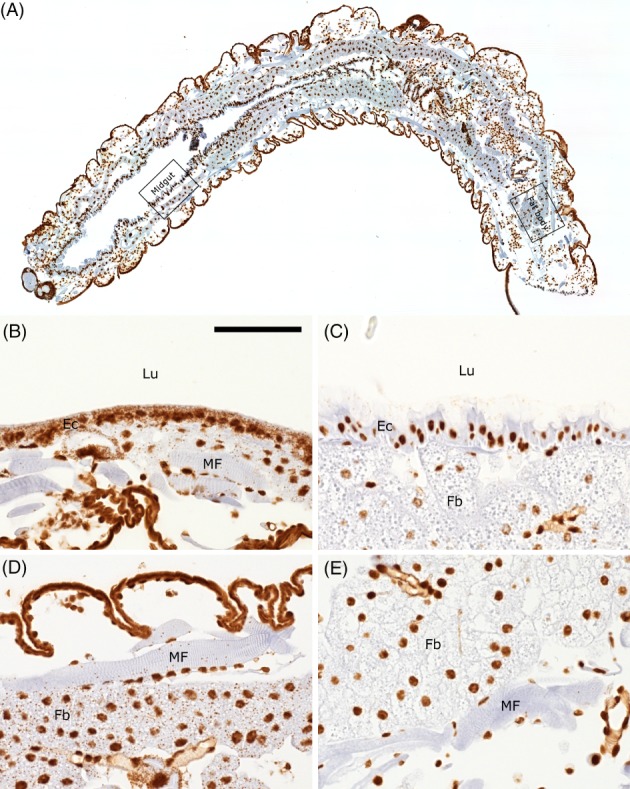
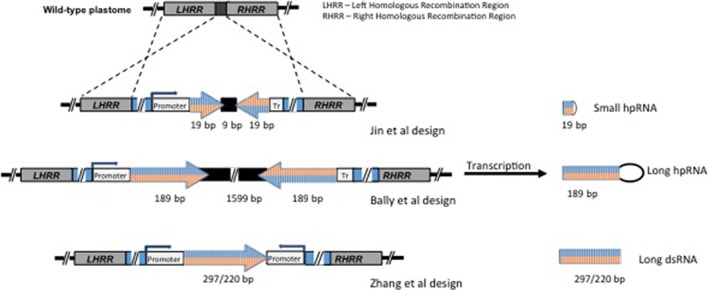
RNA interference (RNAi) was discovered almost 20 years ago and has been exploited worldwide to silence genes in plants and animals. A decade later, it was found that transforming plants with an RNAi construct targeting an insect gene could protect the plant against feeding by that insect. Production of double-stranded RNA (dsRNA) in a plant to affect the viability of a herbivorous animal is termed trans-kingdom RNAi (TK-RNAi). Since this pioneering work, there have been many further examples of successful TK-RNAi, but also reports of failed attempts and unrepeatable experiments. Recently, three laboratories have shown that producing dsRNA in a plant's chloroplast, rather than in its cellular cytoplasm, is a very effective way of delivering TK-RNAi. Our review examines this potentially game-changing approach and compares it with other transgenic insect-proofing schemes. © 2018 The Authors. Pest Management Science published by John Wiley & Sons Ltd on behalf of Society of Chemical Industry.
Keywords: Chloroplast; Kingdom; RNAi; Trans.
© 2018 The Authors. Pest Management Science published by John Wiley & Sons Ltd on behalf of Society of Chemical Industry.
Figures
References
-
- Culliney TW, Crop Losses to Arthropods, in Pimentel D, Peshin R, editors. Integrated Pest Management: Pesticide Problems, Vol 3, Springer Netherlands, Dordrecht, pp. 201–225 (2014).
-
- Narva KE, Storer NP and Meade T, Chapter Four ‐ Discovery and Development of Insect‐Resistant Crops Using Genes from Bacillus thuringiensis, in Advances in Insect Physiology, ed. by Tarlochan SD. and Sarjeet SG. Vol. 47, Academic Press, pp. 177–247 (2014).
-
- Carrière Y, Fabrick JA and Tabashnik BE, Can Pyramids and Seed Mixtures Delay Resistance to Bt Crops? Trends Biotechnol 34:291–302 (2016). - PubMed
-
- Baum JA, Bogaert T, Clinton W, Heck GR, Feldmann P, Ilagan O, et al, Control of coleopteran insect pests through RNA interference. Nature Biotechnol 25:1322–1326 (2007). - PubMed
-
- Mao Y‐B, Cai W‐J, Wang J‐W, Hong G‐J, Tao X‐Y, Wang L‐J et al, Silencing a cotton bollworm P450 monooxygenase gene by plant‐mediated RNAi impairs larval tolerance of gossypol. Nat Biotech 25:1307–1313 (2007). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
