

Rewired RNAi-mediated genome surveillance in house dust mites
- PMID: 29377900
- PMCID: PMC5805368
- DOI: 10.1371/journal.pgen.1007183
Rewired RNAi-mediated genome surveillance in house dust mites
Abstract
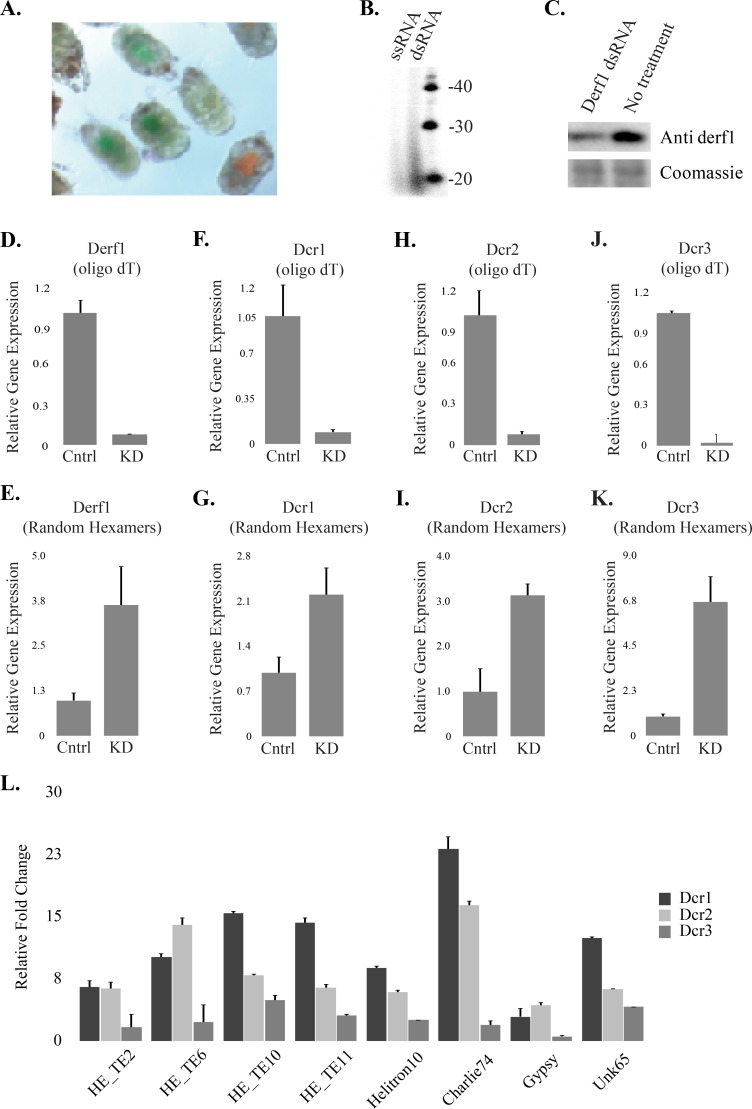
House dust mites are common pests with an unusual evolutionary history, being descendants of a parasitic ancestor. Transition to parasitism is frequently accompanied by genome rearrangements, possibly to accommodate the genetic change needed to access new ecology. Transposable element (TE) activity is a source of genomic instability that can trigger large-scale genomic alterations. Eukaryotes have multiple transposon control mechanisms, one of which is RNA interference (RNAi). Investigation of the dust mite genome failed to identify a major RNAi pathway: the Piwi-associated RNA (piRNA) pathway, which has been replaced by a novel small-interfering RNA (siRNA)-like pathway. Co-opting of piRNA function by dust mite siRNAs is extensive, including establishment of TE control master loci that produce siRNAs. Interestingly, other members of the Acari have piRNAs indicating loss of this mechanism in dust mites is a recent event. Flux of RNAi-mediated control of TEs highlights the unusual arc of dust mite evolution.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





Comment in
-
House dust mites use a plant-like siRNA pathway to silence transposable elements.PLoS Genet. 2018 Apr 5;14(4):e1007255. doi: 10.1371/journal.pgen.1007255. eCollection 2018 Apr. PLoS Genet. 2018. PMID: 29621231 Free PMC article. No abstract available.
References
-
- Arlian LG. House-dust-mite allergens: a review. Exp Appl Acarol. 1991;10(3–4):167–86. . - PubMed
-
- Klimov PB, OC B. Is permanent parasitism reversible?—critical evidence from early evolution of house dust mites. Syst Biol. 2013;62(3):411–23. doi: 10.1093/sysbio/syt008 . - DOI - PubMed
-
- Brookfield JF. Host-parasite relationships in the genome. BMC Biol. 2011;9:67 doi: 10.1186/1741-7007-9-67 ; PubMed Central PMCID: PMCPMC3189907. - DOI - PMC - PubMed
-
- Poulin R, Randhawa HS. Evolution of parasitism along convergent lines: from ecology to genomics. Parasitology. 2015;142 Suppl 1:S6–S15. doi: 10.1017/S0031182013001674 ; PubMed Central PMCID: PMCPMC4413784. - DOI - PMC - PubMed
-
- Fedoroff NV. Presidential address. Transposable elements, epigenetics, and genome evolution. Science. 2012;338(6108):758–67. . - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
