

Roles of Three Cryptococcus neoformans and Cryptococcus gattii Efflux Pump-Coding Genes in Response to Drug Treatment
- PMID: 29378705
- PMCID: PMC5913978
- DOI: 10.1128/AAC.01751-17
Roles of Three Cryptococcus neoformans and Cryptococcus gattii Efflux Pump-Coding Genes in Response to Drug Treatment
Abstract
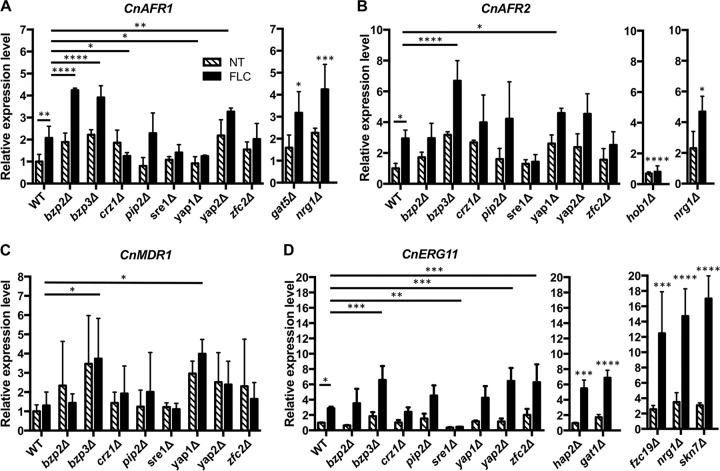
Cryptococcus neoformans and Cryptococcus gattii species complexes are the etiologic agents of cryptococcosis. We have deciphered the roles of three ABC transporters, Afr1, Afr2, and Mdr1, in the representative strains of the two species, C. neoformans H99 and C. gattii R265. Deletion of AFR1 in H99 and R265 drastically reduced the levels of resistance to three xenobiotics and three triazoles, suggesting that Afr1 is the major drug efflux pump in both strains. Fluconazole susceptibility was not affected when AFR2 or MDR1 was deleted in both strains. However, when these genes were deleted in combination with AFR1, a minor additive effect in susceptibility toward several drugs was observed. Deletion of all three genes in both strains caused further increases in susceptibility toward fluconazole and itraconazole, suggesting that Afr2 and Mdr1 augment Afr1 function in pumping these triazoles. Intracellular accumulation of Nile Red significantly increased in afr1Δ mutants of both strains, but rhodamine 6G accumulation increased only in the mdr1Δ mutant of H99. Thus, the three efflux pumps play different roles in the two strains when exposed to different azoles and xenobiotics. AFR1 and AFR2 expression was upregulated in H99 and R265 when treated with fluconazole. However, MDR1 expression was upregulated only in R265 under the same conditions. We screened a library of transcription factor mutants and identified several mutants that manifested either altered fluconazole sensitivity or an increase in the frequency of fluconazole heteroresistance. Gene expression analysis suggests that the three efflux pumps are regulated independently by different transcription factors in response to fluconazole exposure.
Keywords: ABC transport; Afr1; Afr2; Mdr1; azoles; efflux pump.
Copyright © 2018 American Society for Microbiology.
Figures
References
-
- Kwon-Chung KJ, Bennett JE, Wickes BL, Meyer W, Cuomo CA, Wollenburg KR, Bicanic TA, Castaneda E, Chang YC, Chen J, Cogliati M, Dromer F, Ellis D, Filler SG, Fisher MC, Harrison TS, Holland SM, Kohno S, Kronstad JW, Lazera M, Levitz SM, Lionakis MS, May RC, Ngamskulrongroj P, Pappas PG, Perfect JR, Rickerts V, Sorrell TC, Walsh TJ, Williamson PR, Xu J, Zelazny AM, Casadevall A. 2017. The case for adopting the “species complex” nomenclature for the etiologic agents of cryptococcosis. mSphere 2:e00357-16. doi: 10.1128/mSphere.00357-16. - DOI - PMC - PubMed
-
- Perfect JR, Dismukes WE, Dromer F, Goldman DL, Graybill JR, Hamill RJ, Harrison TS, Larsen RA, Lortholary O, Nguyen MH, Pappas PG, Powderly WG, Singh N, Sobel JD, Sorrell TC. 2010. Clinical practice guidelines for the management of cryptococcal disease: 2010 update by the infectious diseases society of america. Clin Infect Dis 50:291–322. doi: 10.1086/649858. - DOI - PMC - PubMed
-
- Norman AW, Demel RA, de Kruyff B, Geurts van Kessel WS, van Deenen LL. 1972. Studies on the biological properties of polyene antibiotics: comparison of other polyenes with filipin in their ability to interact specifically with sterol. Biochim Biophys Acta 290:1–14. doi: 10.1016/0005-2736(72)90046-6. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
