

Immunomodulatory capacity of the serotonin receptor 5-HT2B in a subset of human dendritic cells
- PMID: 29379077
- PMCID: PMC5788853
- DOI: 10.1038/s41598-018-20173-y
Immunomodulatory capacity of the serotonin receptor 5-HT2B in a subset of human dendritic cells
Abstract
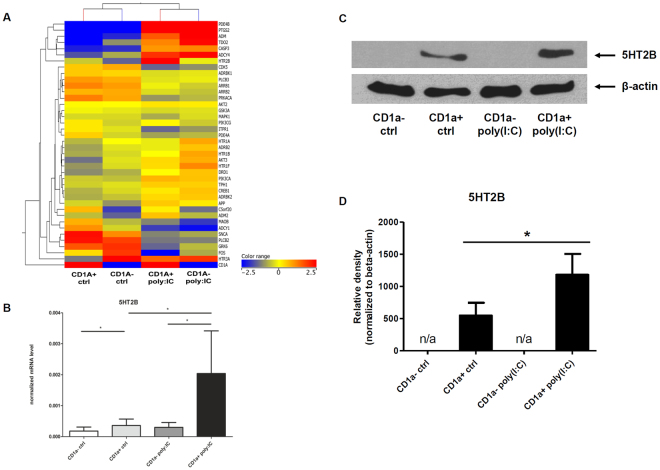
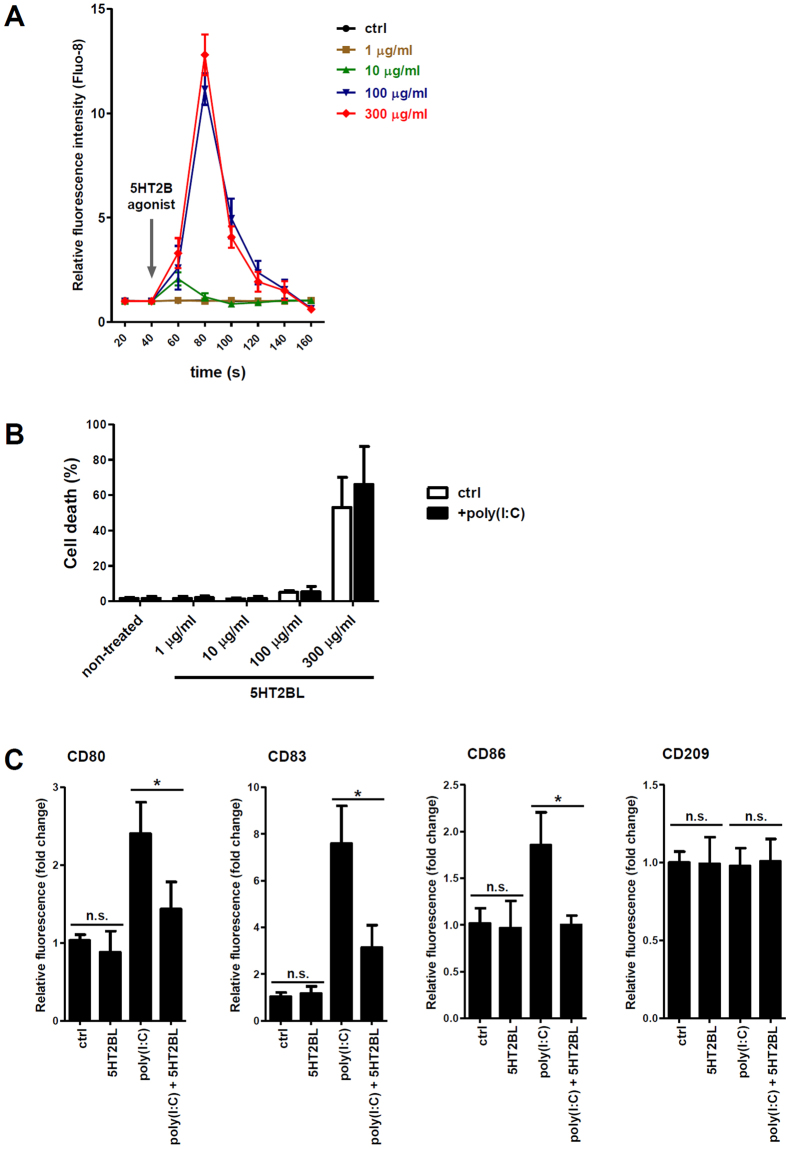
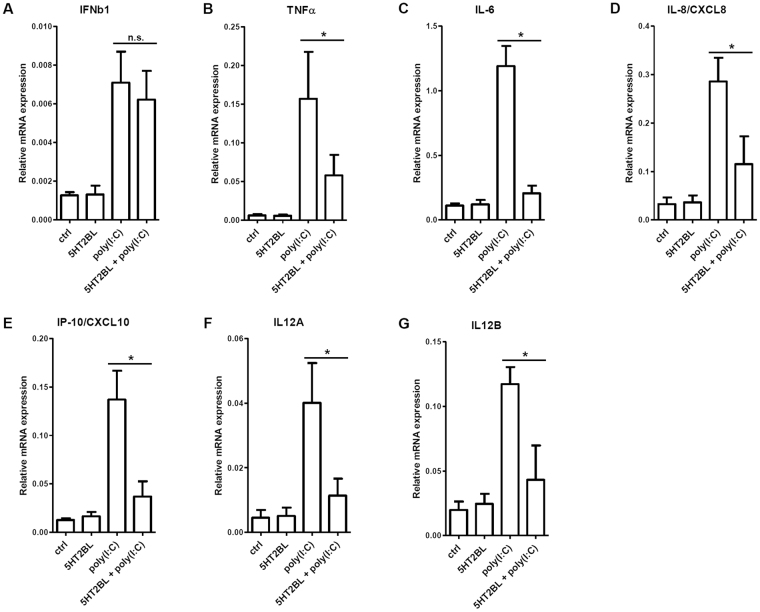
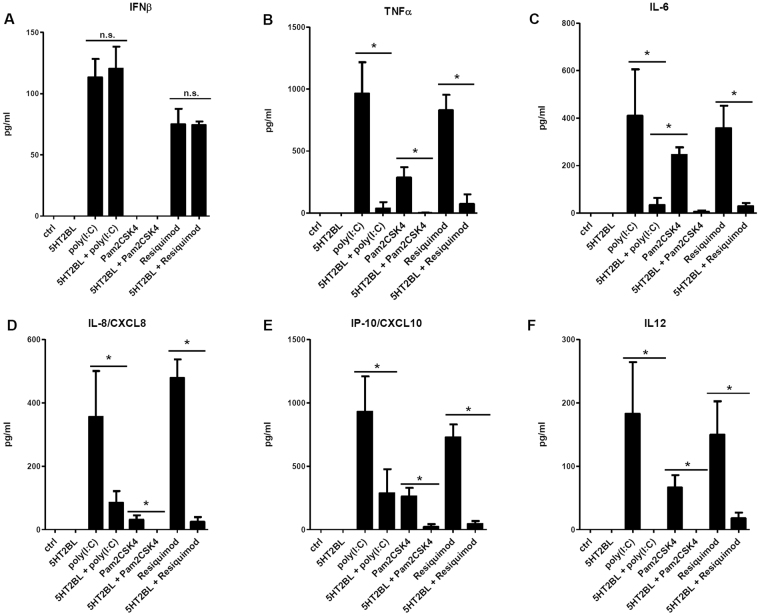
Serotonin is a monoamine neurotransmitter that signals through a wide array of receptors (5-HT1-7) many of which are also involved in immune processes. Dendritic cells (DCs) are crucial players in immune defense by bridging innate and adaptive immune responses via their vast repertoire of pattern recognition receptors and antigen-presenting capability. Although serotonin is known to influence immunity at many levels, cell type-specific expression and function of its receptors remains poorly understood. Here we aimed to study 5-HT1-7 expression and function in CD1a- and CD1a+ human monocyte-derived DCs (moDCs). We found that the 5-HT2B receptor-subtype is solely expressed by the inflammatory CD1a+ moDC subset. Specific 5-HT2B activation potently inhibited TLR2, TLR3, and TLR7/8-induced proinflammatory cytokine and chemokine (TNF-α, IL-6, IL-8, IP-10, IL-12) but not type I interferon-β responses. 5-HT2B agonism also interfered with the polarization of CD1a+ moDC-primed CD4+ T cells towards inflammatory Th1 and Th17 effector lymphocytes. Here we report the subset-specific expression and immunomodulatory function of 5-HT2B in human moDCs. Our results expand the biological role of 5-HT2B which may act not only as a neurotransmitter receptor, but also as an important modulator of both innate and adaptive immune responses.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
